
–Ш—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П Tamotsu: –њ—А–Є–µ–Љ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–∞ —Б –њ–Є—Й–µ–є —Г–≤–µ–ї–Є—З–Є–≤–∞–µ—В —Б–Њ–і–µ—А–ґ–∞–љ–Є–µ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–∞ –≤ –Љ–µ–Љ–±—А–∞–љ–∞—Е—Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤ —Г –Ї—А—Л—Б
12.03.2023
–Я—А–Є–µ–Љ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–∞ —Б –њ–Є—Й–µ–є —Г–≤–µ–ї–Є—З–Є–≤–∞–µ—В —Б–Њ–і–µ—А–ґ–∞–љ–Є–µ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–∞ –≤ –Љ–µ–Љ–±—А–∞–љ–∞—Е —Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤ —Г –Ї—А—Л—Б
Mawatari et al. Lipids in Health and Disease 2012,11:161http://www.lipidworid.com/content/11/1/161
https://pubmed.ncbi.nlm.nih.gov/23170810/
–®–Є—А–Њ –Ь–∞–≤–∞—В–∞—А–Є1*, –Ґ–Њ—Б–Є—Е–Є–Ї–Њ –Ъ–∞—В–∞—Д—Г—З–Є 2, –Ъ–Є–є–Њ—В–∞–Ї–∞ –Ь–Є–∞–Ї–µ3 –Є –Ґ–∞–Ї–µ—Е–Є–Ї–Њ –§—Г–і–Ј–Є–љ–Њ4
* –Ю—В–≤–µ—В—Б—В–≤–µ–љ–љ—Л–є –∞–≤—В–Њ—А: mawatari@rheology.po-jp.com
1–Ш–љ—Б—В–Є—В—Г—В —А–µ–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–є —Д—Г–љ–Ї—Ж–Є–Є –њ–Є—В–∞–љ–Є—П, 2241 Kubara, Hisayama-chou, Kasuya-gun, Fukuoka 811-2501, Japan
–Я–Њ–ї–љ—Л–µ —Б–≤–µ–і–µ–љ–Є—П –Њ–± –∞–≤—В–Њ—А–∞—Е –њ—А–Є–≤–µ–і–µ–љ—Л –≤ –Ї–Њ–љ—Ж–µ —Б—В–∞—В—М–Є.
\u00a9 2012 Mawatari et al.; –ї–Є—Ж–µ–љ–Ј–Є–∞—В BioMed Central Ltd. –Э–∞—Б—В–Њ—П—Й–∞—П —Б—В–∞—В—М—П –љ–∞—Е–Њ–і–Є—В—Б—П –≤ –Њ—В–Ї—А—Л—В–Њ–Љ –і–Њ—Б—В—Г–њ–µ –≤ —Б–Њ–Њ—В–≤–µ—В—Б—В–≤–Є–Є —Б —Г—Б–ї–Њ–≤–Є—П–Љ–Є –ї–Є—Ж–µ–љ–Ј–Є–Є Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0~~HEAD=pobj), —А–∞–Ј—А–µ—И–∞—О—Й–µ–є –љ–µ–Њ–≥—А–∞–љ–Є—З–µ–љ–љ–Њ–µ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–µ, —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–Є–µ –Є –≤–Њ—Б–њ—А–Њ–Є–Ј–≤–µ–і–µ–љ–Є–µ –љ–∞ –ї—О–±–Њ–Љ –љ–Њ—Б–Є—В–µ–ї–µ –њ—А–Є —Г—Б–ї–Њ–≤–Є–Є –њ—А–∞–≤–Є–ї—М–љ–Њ–≥–Њ —Ж–Є—В–Є—А–Њ–≤–∞–љ–Є—П –Њ—А–Є–≥–Є–љ–∞–ї—М–љ–Њ–є —А–∞–±–Њ—В—Л.
–Р–≤—В–Њ—А–µ—Д–µ—А–∞—В
–Т–≤–µ–і–µ–љ–Є–µ: –Є–Ј–≤–µ—Б—В–љ–Њ –Њ –Љ–љ–Њ–≥–Є—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П—Е, —Б–≤—П–Ј–∞–љ–љ—Л—Е —Б –і–µ—Д–Є—Ж–Є—В–Њ–Љ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤. –£–≤–µ–ї–Є—З–µ–љ–Є–µ —Б–Њ–і–µ—А–ґ–∞–љ–Є—П –Є–ї–Є –Ј–∞–Љ–µ—Й–µ–љ–Є–µ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ –≤ —В–Ї–∞–љ—П—Е –Љ–Њ–≥–ї–Њ –±—Л –±–ї–∞–≥–Њ—В–≤–Њ—А–љ–Њ –≤–ї–Є—П—В—М –љ–∞ —Б–Њ—Б—В–Њ—П–љ–Є–µ –њ–∞—Ж–Є–µ–љ—В–Њ–≤, —Б—В—А–∞–і–∞—О—Й–Є—Е –Њ—В —В–∞–Ї–Є—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є–є, –Њ–і–љ–∞–Ї–Њ –Є–љ—Д–Њ—А–Љ–∞—Ж–Є—П –Њ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є–Є –њ–Є—Й–µ–≤—Л—Е –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ –љ–∞ –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –Њ—В—Б—Г—В—Б—В–≤—Г–µ—В.
–Ь–µ—В–Њ–і: –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л –±—Л–ї–Є –њ–Њ–ї—Г—З–µ–љ—Л –Є–Ј –Ї—Г—А–Є–љ–Њ–є –Ї–Њ–ґ–Є. –Ю—З–Є—Й–µ–љ–љ—Л–µ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л —Б–Њ–і–µ—А–ґ–∞–ї–Є 96,4% —Н—В–∞–љ–Њ–ї–∞–Љ–Є–љ-–њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–∞ (PlsEtn), 2,4% —Е–Њ–ї–Є–љ-–њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–∞ (PlsCho) –Є 0,5% —Б—Д–Є–љ–≥–Њ–Љ–Є–µ–ї–Є–љ–∞ (SM).
–Ъ—А—Л—Б–∞–Љ –±—Л–ї–∞ –њ—А–µ–і–ї–Њ–ґ–µ–љ–∞ –і–Є–µ—В–∞, —Б–Њ–і–µ—А–ґ–∞—Й–∞—П 0,1% –Њ—З–Є—Й–µ–љ–љ—Л—Е –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ (–і–Є–µ—В–∞ PlsEtn). –Ф–ї—П –Њ–њ—А–µ–і–µ–ї–µ–љ–Є—П –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ–≥–Њ —Б–Њ–і–µ—А–ґ–∞–љ–Є—П —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤ –±—Л–ї –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ –Љ–µ—В–Њ–і –≤—Л—Б–Њ–Ї–Њ—Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ–є –ґ–Є–і–Ї–Њ—Б—В–љ–Њ–є —Е—А–Њ–Љ–∞—В–Њ–≥—А–∞—Д–Є–Є (–Т–≠–Ц–•), –њ–Њ–Ј–≤–Њ–ї—П—О—Й–Є–є —А–∞–Ј–і–µ–ї—П—В—М –Є–љ—В–∞–Ї—В–љ—Л–µ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л –Є –≤—Б–µ –і—А—Г–≥–Є–µ –Ї–ї–∞—Б—Б—Л —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤ –Ј–∞ –Њ–і–Є–љ —Е—А–Њ–Љ–∞—В–Њ–≥—А–∞—Д–Є—З–µ—Б–Ї–Є–є –њ—А–Њ–≥–Њ–љ.
–†–µ–Ј—Г–ї—М—В–∞—В—Л: —Б–ї–µ–і—Б—В–≤–Є–µ–Љ –њ—А–Є–µ–Љ–∞ –і–Є–µ—В—Л PlsEtn, –Ї–Њ—В–Њ—А—Г—О –Ї—А—Л—Б—Л –¶—Г–Ї–µ—А, —Б—В—А–∞–і–∞—О—Й–Є–µ –і–Є–∞–±–µ—В–Њ–Љ –Є –Њ–ґ–Є—А–µ–љ–Є–µ–Љ (ZDF), –њ–Њ–ї—Г—З–∞–ї–Є –≤ —В–µ—З–µ–љ–Є–µ 4 –љ–µ–і–µ–ї—М, —Б—В–∞–ї–Њ —Б–љ–Є–ґ–µ–љ–Є–µ —Г—А–Њ–≤–љ—П —Е–Њ–ї–µ—Б—В–µ—А–Є–љ–∞ –Є —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤ –≤ –њ–ї–∞–Ј–Љ–µ –њ–Њ —Б—А–∞–≤–љ–µ–љ–Є—О —Б –Ї–Њ–љ—В—А–Њ–ї—М–љ–Њ–є –і–Є–µ—В–Њ–є.
–†–µ–Ј—Г–ї—М—В–∞—В—Л –і—А—Г–≥–Є—Е —Б—В–∞–љ–і–∞—А—В–љ—Л—Е –ї–∞–±–Њ—А–∞—В–Њ—А–љ—Л—Е –∞–љ–∞–ї–Є–Ј–Њ–≤ –њ–ї–∞–Ј–Љ—Л, –≤ —В–Њ–Љ —З–Є—Б–ї–µ –љ–∞ —В—А–Є–≥–ї–Є—Ж–µ—А–Є–і –Є –≥–ї—О–Ї–Њ–Ј—Г, –њ–µ—З–µ–љ–Њ—З–љ–∞—П –Є –њ–Њ—З–µ—З–љ–∞—П —Д—Г–љ–Ї—Ж–Є—П, —Г—А–Њ–≤–µ–љ—М –∞–ї—М–±—Г–Љ–Є–љ–∞ –Є –Љ–∞—Б—Б–∞ —В–µ–ї–∞ –љ–µ –њ–Њ–Ї–∞–Ј–∞–ї–Є –Є–Ј–Љ–µ–љ–µ–љ–Є–є. –Т –≥—А—Г–њ–њ–µ, –њ–Њ–ї—Г—З–∞–≤—И–µ–є –і–Є–µ—В—Г PlsEtn, –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ–µ —Б–Њ–і–µ—А–ґ–∞–љ–Є–µ PlsEtn —Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤ –Є —Д–Њ—Б—Д–∞—В–Є–і–Є–ї—Н—В–∞–љ–Њ–ї–∞–Љ–Є–љ–∞ (PE) —Г–≤–µ–ї–Є—З–Є–ї–Њ—Б—М, –∞ –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ–µ —Б–Њ–і–µ—А–ґ–∞–љ–Є–µ —Д–Њ—Б—Д–∞—В–Є–і–Є–ї—Е–Њ–ї–Є–љ–∞ (PC) —Г–Љ–µ–љ—М—И–Є–ї–Њ—Б—М.
–Я—А–Є–µ–Љ –і–Є–µ—В—Л PlsEtn –Ј–і–Њ—А–Њ–≤—Л–Љ–Є –Ї—А—Л—Б–∞–Љ–Є –≤ —В–µ—З–µ–љ–Є–µ 9 –љ–µ–і–µ–ї—М —В–∞–Ї–ґ–µ –њ—А–Є–≤–µ–ї –Ї —Б–љ–Є–ґ–µ–љ–Є—О —Г—А–Њ–≤–љ—П —Е–Њ–ї–µ—Б—В–µ—А–Є–љ–∞ –Є —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤ –≤ –њ–ї–∞–Ј–Љ–µ –Є –≤—Л–Ј–≤–∞–ї —Г–≤–µ–ї–Є—З–µ–љ–Є–µ –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ–≥–Њ —Б–Њ–і–µ—А–ґ–∞–љ–Є—П PlsEtn –≤ –Љ–µ–Љ–±—А–∞–љ–µ —Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤. –†–µ–Ј—Г–ї—М—В–∞—В—Л –і—А—Г–≥–Є—Е —Б—В–∞–љ–і–∞—А—В–љ—Л—Е –ї–∞–±–Њ—А–∞—В–Њ—А–љ—Л—Е –∞–љ–∞–ї–Є–Ј–Њ–≤ –њ–ї–∞–Ј–Љ—Л –Є –Љ–∞—Б—Б–∞ —В–µ–ї–∞ –љ–µ –њ–Њ–Ї–∞–Ј–∞–ї–Є –Є–Ј–Љ–µ–љ–µ–љ–Є–є.
–Т—Л–≤–Њ–і—Л: –њ—А–Є–µ–Љ –њ–Є—Й–µ–≤–Њ–≥–Њ PlsEtn —Г–≤–µ–ї–Є—З–Є–≤–∞–µ—В –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ–µ —Б–Њ–і–µ—А–ґ–∞–љ–Є–µ PlsEtn –≤ –Љ–µ–Љ–±—А–∞–љ–µ —Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤ —Г –Ј–і–Њ—А–Њ–≤—Л—Е –Ї—А—Л—Б –Є –Ї—А—Л—Б ZDF –Є —Б–љ–Є–ґ–∞–µ—В —Г—А–Њ–≤–µ–љ—М —Е–Њ–ї–µ—Б—В–µ—А–Є–љ–∞ –Є —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤ –≤ –њ–ї–∞–Ј–Љ–µ. –Я—А–Є–µ–Љ –њ–Є—Й–µ–≤–Њ–≥–Њ PlsEtn –≤ —В–µ—З–µ–љ–Є–µ 9 –љ–µ–і–µ–ї—М –љ–µ –Њ–Ї–∞–Ј–∞–ї –љ–µ–≥–∞—В–Є–≤–љ–Њ–≥–Њ –≤–ї–Є—П–љ–Є—П –љ–∞ —Б–Њ—Б—В–Њ—П–љ–Є–µ –Ј–і–Њ—А–Њ–≤—Л—Е –Ї—А—Л—Б.
–Ъ–ї—О—З–µ–≤—Л–µ —Б–ї–Њ–≤–∞: –њ–Є—Й–µ–≤–Њ–є –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ, –Ї—А—Л—Б–∞ –¶—Г–Ї–µ—А, —Б—В—А–∞–і–∞—О—Й–∞—П –і–Є–∞–±–µ—В–Њ–Љ –Є –Њ–ґ–Є—А–µ–љ–Є–µ–Љ, –Ї—А—Л—Б–∞ –Т–Є—Б—В–∞—А, —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і—Л —Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤, —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і—Л –њ–ї–∞–Ј–Љ—Л
–Т–≤–µ–і–µ–љ–Є–µ
–Я–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л —П–≤–ї—П—О—В—Б—П –≥–ї–Є—Ж–µ—А–Њ—Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–∞–Љ–Є, –і–ї—П –Ї–Њ—В–Њ—А—Л—Е —Е–∞—А–∞–Ї—В–µ—А–љ–∞ –≤–Є–љ–Є–ї—Н—Д–Є—А–љ–∞—П —Б–≤—П–Ј—М –≤ –њ–Њ–ї–Њ–ґ–µ–љ–Є–Є sn-1 –Њ—Б–љ–Њ–≤–љ–Њ–є —Ж–µ–њ–Є –≥–ї–Є—Ж–µ—А–Њ–ї–∞ –Є —Н—Д–Є—А–љ–∞—П —Б–≤—П–Ј—М –≤ –њ–Њ–ї–Њ–ґ–µ–љ–Є–Є sn-2. –Я–Њ–ї–Њ–ґ–µ–љ–Є–µ sn-1 —З–∞—Й–µ –≤—Б–µ–≥–Њ —Б–≤—П–Ј–∞–љ–Њ —Б –ґ–Є—А–љ—Л–Љ–Є —Б–њ–Є—А—В–∞–Љ–Є C16, –°18, –Є–ї–Є –°18:1, –∞ –њ–Њ–ї–Њ–ґ–µ–љ–Є–µ sn-2, –Ї–∞–Ї –њ—А–∞–≤–Є–ї–Њ, –Ј–∞–љ–Є–Љ–∞–µ—В –њ–Њ–ї–Є–љ–µ–љ–∞—Б—Л—Й–µ–љ–љ–∞—П –ґ–Є—А–љ–∞—П –Ї–Є—Б–ї–Њ—В–∞, –≤ —З–∞—Б—В–љ–Њ—Б—В–Є, –∞—А–∞—Е–Є–і–Њ–љ–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ (ARA) –Є–ї–Є –і–Њ–Ї–Њ–Ј–∞–≥–µ–Ї—Б–∞–µ–љ–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ (DHA) [1-5].–Я–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л –њ—А–Є—Б—Г—В—Б—В–≤—Г—О—В –њ–Њ—З—В–Є –≤–Њ –≤—Б–µ—Е —В–Ї–∞–љ—П—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –Є —Б–Њ—Б—В–∞–≤–ї—П—О—В –Њ–Ї–Њ–ї–Њ 18% –Њ—В –Њ–±—Й–µ–≥–Њ –Њ–±—К–µ–Љ–∞ —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤ –≤ –Ї–ї–µ—В–Њ—З–љ—Л—Е –Љ–µ–Љ–±—А–∞–љ–∞—Е, –њ—А–Є—З–µ–Љ —Н—В–∞–љ–Њ–ї–∞–Љ–Є–љ-–њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л (PlsEtn) –≤—Б—В—А–µ—З–∞—О—В—Б—П –љ–∞–Љ–љ–Њ–≥–Њ —З–∞—Й–µ, —З–µ–Љ —Е–Њ–ї–Є–љ-–њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л (PlsCho), –Ј–∞ –Є—Б–Ї–ї—О—З–µ–љ–Є–µ–Љ —Б–µ—А–і–µ—З–љ–Њ–є –Є —Б–Ї–µ–ї–µ—В–љ–Њ–є –Љ—Л—И–µ—З–љ–Њ–є —В–Ї–∞–љ–Є [1-5]. –Я–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л –љ–µ —В–Њ–ї—М–Ї–Њ —П–≤–ї—П—О—В—Б—П —Б—В—А—Г–Ї—В—Г—А–љ—Л–Љ –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В–Њ–Љ –Љ–µ–Љ–±—А–∞–љ—Л –Є —Е—А–∞–љ–Є–ї–Є—Й–µ–Љ –≤—В–Њ—А–Є—З–љ—Л—Е –њ–Њ—Б—А–µ–і–љ–Є–Ї–Њ–≤ —Г –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е, –љ–Њ –Є –Љ–Њ–≥—Г—В —Г—З–∞—Б—В–≤–Њ–≤–∞—В—М –≤ –њ—А–Њ—Ж–µ—Б—Б–∞—Е –Љ–µ–Љ–±—А–∞–љ–љ–Њ–≥–Њ —Б–Є–љ—В–µ–Ј–∞, –њ–µ—А–µ–љ–Њ—Б–∞ –Є–Њ–љ–Њ–≤ –Є –Њ—В—В–Њ–Ї–∞ —Е–Њ–ї–µ—Б—В–µ—А–Є–љ–∞ [1-5].
–Т–Є–љ–Є–ї—Н—Д–Є—А–љ–∞—П —Б–≤—П–Ј—М –≤ –њ–Њ–ї–Њ–ґ–µ–љ–Є–Є sn-1 –і–µ–ї–∞–µ—В –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л –±–Њ–ї–µ–µ –≤–Њ—Б–њ—А–Є–Є–Љ—З–Є–≤—Л–Љ–Є –Ї –Њ–Ї–Є—Б–ї–Є—В–µ–ї—М–љ–Њ–Љ—Г —Б—В—А–µ—Б—Б—Г –њ–Њ —Б—А–∞–≤–љ–µ–љ–Є—О —Б —Б–Њ–Њ—В–≤–µ—В—Б—В–≤—Г—О—Й–Є–Љ–Є –≥–ї–Є—Ж–µ—А–Њ—Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–∞–Љ–Є —Б —Н—Д–Є—А–љ—Л–Љ–Є —Б–≤—П–Ј—П–Љ–Є. –Я–Њ—Н—В–Њ–Љ—Г –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л –Љ–Њ–≥—Г—В —В–∞–Ї–ґ–µ –≤—Л—Б—В—Г–њ–∞—В—М –≤ –Ї–∞—З–µ—Б—В–≤–µ –∞–љ—В–Є–Њ–Ї—Б–Є–і–∞–љ—В–Њ–≤, –Ј–∞—Й–Є—Й–∞—О—Й–Є—Е –Ї–ї–µ—В–Ї–Є –Њ—В –Њ–Ї–Є—Б–ї–Є—В–µ–ї—М–љ–Њ–≥–Њ —Б—В—А–µ—Б—Б–∞ [1-5].
–С–Є–Њ—Б–Є–љ—В–µ–Ј –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ –љ–∞—З–Є–љ–∞–µ—В—Б—П –≤ –њ–µ—А–Њ–Ї—Б–Є—Б–Њ–Љ–∞—Е, –Є –≤—Б—В—А–µ—З–∞—О—Й–∞—П—Б—П —Г –ї—О–і–µ–є –љ–∞—Б–ї–µ–і—Б—В–≤–µ–љ–љ–∞—П –њ–µ—А–Њ–Ї—Б–Є—Б–Њ–Љ–љ–∞—П –њ–∞—В–Њ–ї–Њ–≥–Є—П, —А–Є–Ј–Њ–Љ–µ–ї–Є—З–µ—Б–Ї–∞—П —В–Њ—З–µ—З–љ–∞—П —Е–Њ–љ–і—А–Њ–і–Є—Б–њ–ї–∞–Ј–Є—П (RCDP), –њ—А–Њ—П–≤–ї—П–µ—В—Б—П –≤ –і–µ—Д–Є—Ж–Є—В–µ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ —В–Ї–∞–љ–µ–є —А–∞–Ј–ї–Є—З–љ—Л—Е –Њ—А–≥–∞–љ–Њ–≤ –Є –≤—Л–Ј—Л–≤–∞–µ—В —Б–µ—А—М–µ–Ј–љ—Л–µ —А–∞—Б—Б—В—А–Њ–є—Б—В–≤–∞ –Љ–љ–Њ–≥–Є—Е —В–Ї–∞–љ–µ–є –Є –Њ—А–≥–∞–љ–Њ–≤, —В–∞–Ї–Є—Е –Ї–∞–Ї —Б–Ї–µ–ї–µ—В, –Љ–Њ–Ј–≥, —Е—А—Г—Б—В–∞–ї–Є–Ї –≥–ї–∞–Ј–∞, –њ–Њ—З–Ї–Є –Є —Б–µ—А–і—Ж–µ [2-5]. RCDP –Љ–Њ–ґ–µ—В –±—Л—В—М –њ—А—П–Љ—Л–Љ —Б–ї–µ–і—Б—В–≤–Є–µ–Љ –і–µ—Д–Є—Ж–Є—В–∞ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ –Є —Г–Ї–∞–Ј—Л–≤–∞–µ—В –љ–∞ –≤–∞–ґ–љ–Њ—Б—В—М –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ –≤ –Њ—А–≥–∞–љ–Є–Ј–Љ–µ —З–µ–ї–Њ–≤–µ–Ї–∞.
–Т —В–Њ –ґ–µ –≤—А–µ–Љ—П, –≤—В–Њ—А–Є—З–љ—Л–є –і–µ—Д–Є—Ж–Є—В –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ –љ–∞–±–ї—О–і–∞–µ—В—Б—П –њ—А–Є –Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Є—Е –Є –≤–Њ—Б–њ–∞–ї–Є—В–µ–ї—М–љ—Л—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П—Е, —В–∞–Ї–Є—Е –Ї–∞–Ї —Б–∞—Е–∞—А–љ—Л–є –і–Є–∞–±–µ—В, —Б–µ—А–і–µ—З–љ—Л–µ –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П, —А–∞–Ї, —А–µ—Б–њ–Є—А–∞—В–Њ—А–љ—Л–µ –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П –Є –±–Њ–ї–µ–Ј–љ—М –Р–ї—М—Ж–≥–µ–є–Љ–µ—А–∞ [3,4]. –Т—В–Њ—А–Є—З–љ—Л–є –і–µ—Д–Є—Ж–Є—В –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ –Љ–Њ–ґ–µ—В –±—Л—В—М —Б–ї–µ–і—Б—В–≤–Є–µ–Љ –Ј–∞–Љ–µ–і–ї–µ–љ–Є—П —Б–Є–љ—В–µ–Ј–∞ –Є/–Є–ї–Є —Г—Б–Ї–Њ—А–µ–љ–Є—П —А–∞—Б–њ–∞–і–∞ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤. –Я–Њ–њ–Њ–ї–љ–µ–љ–Є–µ –Є/–Є–ї–Є –Ј–∞–Љ–µ—Й–µ–љ–Є–µ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ –Љ–Њ–≥–ї–Њ –±—Л –њ—А–Є–љ–µ—Б—В–Є —Б—Г—Й–µ—Б—В–≤–µ–љ–љ—Г—О –њ–Њ–ї—М–Ј—Г –њ–∞—Ж–Є–µ–љ—В–∞–Љ, —Б—В—А–∞–і–∞—О—Й–Є–Љ —Н—В–Є–Љ–Є –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П–Љ–Є –њ—А–Є –і–µ—Д–Є—Ж–Є—В–µ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤.
–Э–µ–і–∞–≤–љ–Њ –±—Л–ї –≤—Л–њ–Њ–ї–љ–µ–љ –∞–љ–∞–ї–Є–Ј –і–∞–љ–љ—Л—Е –Њ –≤–ї–Є—П–љ–Є–Є –њ–Є—Й–µ–≤—Л—Е —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤ –љ–∞ –Ј–і–Њ—А–Њ–≤—М–µ [6], –Ї–Њ—В–Њ—А—Л–є –њ–Њ–Ї–∞–Ј–∞–ї, —З—В–Њ —Б–Њ–≥–ї–∞—Б–љ–Њ –±–Њ–ї—М—И–Є–љ—Б—В–≤—Г –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є –њ–Є—Й–µ–≤—Л–µ —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і—Л –Њ–Ї–∞–Ј—Л–≤–∞—О—В –њ–Њ–ї–Њ–ґ–Є—В–µ–ї—М–љ–Њ–µ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є–µ –њ—А–Є –љ–µ–Ї–Њ—В–Њ—А—Л—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П—Е –±–µ–Ј –Ї–∞–Ї–Є—Е-–ї–Є–±–Њ —Б–µ—А—М–µ–Ј–љ—Л—Е –њ–Њ–±–Њ—З–љ—Л—Е —Н—Д—Д–µ–Ї—В–Њ–≤. –Ю–і–љ–∞–Ї–Њ –і–Є–µ—В–Є—З–µ—Б–Ї–Є–µ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л –љ–µ –±—Л–ї–Є —Г–њ–Њ–Љ—П–љ—Г—В—Л [6].
–Я–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л –±—Л–ї–Є –њ–Њ–ї—Г—З–µ–љ—Л –љ–∞–Љ–Є –Є–Ј –Ї—Г—А–Є–љ–Њ–є –Ї–Њ–ґ–Є. –Ь—Л –њ—А–µ–і–ї–∞–≥–∞–ї–Є –і–Є–µ—В—Г, —Б–Њ–і–µ—А–ґ–∞—Й—Г—О 0,1% –Њ—З–Є—Й–µ–љ–љ—Л—Е –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ –Ї—А—Л—Б–∞–Љ –¶—Г–Ї–µ—А, —Б—В—А–∞–і–∞—О—Й–Є–Љ –і–Є–∞–±–µ—В–Њ–Љ –Є –Њ–ґ–Є—А–µ–љ–Є–µ–Љ (ZDF), –Є –Ј–і–Њ—А–Њ–≤—Л–Љ –Ї—А—Л—Б–∞–Љ –ї–Є–љ–Є–Є –Т–Є—Б—В–∞—А. –С—Л–ї–Є –≤—Л–њ–Њ–ї–љ–µ–љ—Л –Њ–±—Л—З–љ—Л–µ –ї–∞–±–Њ—А–∞—В–Њ—А–љ—Л–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –њ–ї–∞–Ј–Љ—Л –Ї—А–Њ–≤–Є (–≤ —В–Њ–Љ —З–Є—Б–ї–µ, –љ–∞ –Њ–±—Й–µ–µ —Б–Њ–і–µ—А–ґ–∞–љ–Є–µ —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤) –Є –∞–љ–∞–ї–Є–Ј —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–љ–Њ–≥–Њ —Б–Њ—Б—В–∞–≤–∞ –њ–ї–∞–Ј–Љ—Л –Є —Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤.
–†–µ–Ј—Г–ї—М—В–∞—В—Л –Є –∞–љ–∞–ї–Є–Ј
–Ю—З–Є—Й–µ–љ–љ—Л–µ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л, –њ–Њ–ї—Г—З–µ–љ–љ—Л–µ –Є–Ј –Ї—Г—А–Є–љ–Њ–є –Ї–Њ–ґ–Є, —Б–Њ—Б—В–Њ—П–ї–Є, —Б–Њ–≥–ї–∞—Б–љ–Њ —А–∞—Б—З–µ—В—Г –њ–Њ —Е—А–Њ–Љ–∞—В–Њ–≥—А–∞—Д–Є—З–µ—Б–Ї–Њ–є –Њ–±–ї–∞—Б—В–Є –Т–≠–Ц–•, –Є–Ј 96,4% —Н—В–∞–љ–Њ–ї–∞–Љ–Є–љ-–њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ (PlsEtn), 2,6% —Е–Њ–ї–Є–љ-–њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ (PlsCho), 0,5% SM –Є 0,7% –і—А—Г–≥–Є—Е —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤ (—В–∞–±–ї–Є—Ж–∞ 1, —А–Є—Б. 1).–Т—А–µ–Љ—П —Г–і–µ—А–ґ–Є–≤–∞–љ–Є—П —Е—А–Њ–Љ–∞—В–Њ–≥—А–∞—Д–Є—З–µ—Б–Ї–Є—Е –њ–Є–Ї–Њ–≤ PlsEtn –Є PlsCho —Б–Њ–Њ—В–≤–µ—В—Б—В–≤–Њ–≤–∞–ї–Њ –≤—А–µ–Љ–µ–љ–Є —Г–і–µ—А–ґ–Є–≤–∞–љ–Є—П PlsEtn –Є PlsCho —Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤ –Љ–µ–Љ–±—А–∞–љ—Л –Ї—А—Л—Б—Л (—А–Є—Б. 1). –Я–Є–Ї–Є –Є—Б—З–µ–Ј–∞–ї–Є –њ–Њ—Б–ї–µ –Њ–±—А–∞–±–Њ—В–Ї–Є —Б–Њ–ї—П–љ–Њ–є –Ї–Є—Б–ї–Њ—В–Њ–є (–Э–°l), —З—В–Њ –Њ–Ј–љ–∞—З–∞–µ—В, —З—В–Њ –њ–Є–Ї–Є —Б–Њ–Њ—В–≤–µ—В—Б—В–≤–Њ–≤–∞–ї–Є –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–∞–Љ [7,8] (–і–∞–љ–љ—Л–µ –љ–µ –њ–Њ–Ї–∞–Ј–∞–љ—Л). –Ц–Є—А–љ—Л–µ –Ї–Є—Б–ї–Њ—В—Л PlsEtn —Б–Њ—Б—В–Њ—П–ї–Є –Є–Ј DHA (22:6), ARA (20:4), –Њ–ї–µ–Є–љ–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л (18:1), –Љ–Є—А–Є—Б—В–Є–љ–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л (14:0), –њ–∞–ї—М–Љ–Є—В–Є–љ–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л (16:0) –Є —Б—В–µ–∞—А–Є–љ–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л (18:0) (–Ґ–∞–±–ї–Є—Ж–∞ 1), —З—В–Њ —Б–Њ–Њ—В–≤–µ—В—Б—В–≤—Г–µ—В —Б–Њ—Б—В–∞–≤—Г –Њ–±—Л—З–љ—Л—Е –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ –≤ —В–Ї–∞–љ—П—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е [1-4].
–Ґ–∞–±–ї–Є—Ж–∞ 1 –§–Њ—Б—Д–Њ–ї–Є–њ–Є–і–љ—Л–є —Б–Њ—Б—В–∞–≤ –Њ—З–Є—Й–µ–љ–љ—Л—Е –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ –Є –ґ–Є—А–љ–Њ–Ї–Є—Б–ї–Њ—В–љ—Л–є —Б–Њ—Б—В–∞–≤ PlsEtn –≤ –Њ—З–Є—Й–µ–љ–љ—Л—Е –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–∞—Е
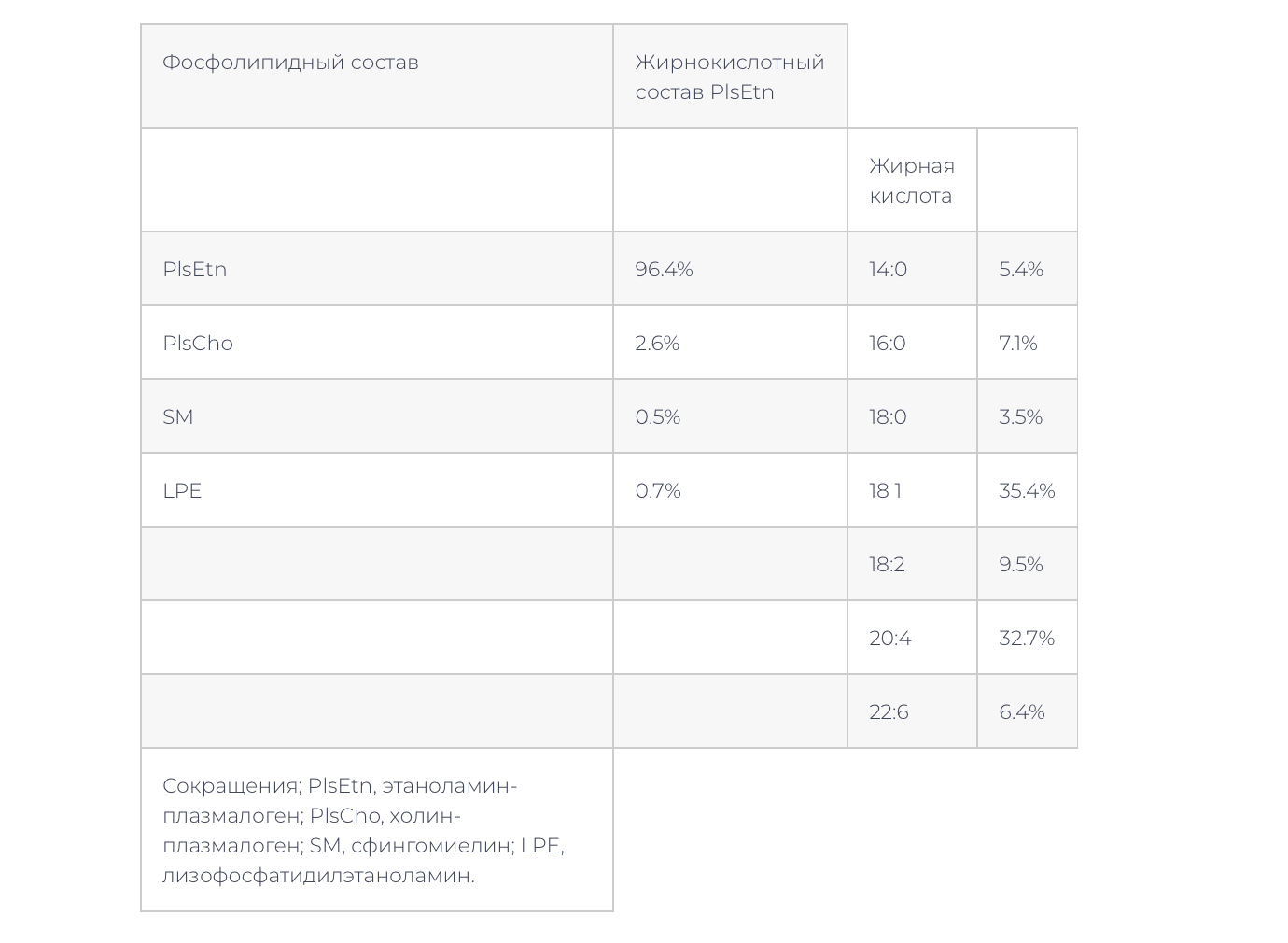
–Ю—З–Є—Й–µ–љ–љ—Л–µ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л
–†–Є—Б—Г–љ–Њ–Ї 1 –Т–≠–Ц–• —Е—А–Њ–Љ–∞—В–Њ–≥—А–∞–Љ–Љ–∞ –Њ—З–Є—Й–µ–љ–љ–Њ–≥–Њ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–∞ –Є–Ј –Ї—Г—А–Є–љ–Њ–є –Ї–Њ–ґ–Є. –Т—А–µ–Љ—П —Г–і–µ—А–ґ–Є–≤–∞–љ–Є—П —Н—В–∞–љ–Њ–ї–∞–Љ–Є–љ-–њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ (PlsEtn) –Є —Е–Њ–ї–Є–љ-–њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ (PlsCho) —Б–Њ–Њ—В–≤–µ—В—Б—В–≤–Њ–≤–∞–ї–Њ –њ–Њ–Ї–∞–Ј—В–µ–ї—П–Љ —Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤ –Ї—А—Л—Б—Л. –Ф—А—Г–≥–Є–µ —Б–Њ–Ї—А–∞—Й–µ–љ–Є—П: PE, —Д–Њ—Б—Д–∞—В–Є–і–Є–ї—Н—В–∞–љ–Њ–ї–∞–Љ–Є–љ; PC, —Д–Њ—Б—Д–∞—В–Є–і–Є–ї—Е–Њ–ї–Є–љ; SM-1 –Є SM-2, —Б—Д–Є–љ–≥–Њ–Љ–Є–µ–ї–Є–љ; PS, —Д–Њ—Б—Д–∞—В–Є–і–Є–ї—Б–µ—А–Є–љ; PI, —Д–Њ—Б—Д–∞—В–Є–і–Є–ї–Є–љ–Њ–Ј–Є—В–Њ–ї.(–Ъ–Њ–љ—В—А–Њ–ї—М–љ–∞—П –і–Є–µ—В–∞)
(–Ф–Є–µ—В–∞ PlsEtn)
–†–Є—Б—Г–љ–Њ–Ї 2 –Т–≠–Ц–•-—Е—А–Њ–Љ–∞—В–Њ–≥—А–∞–Љ–Љ—Л —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤ –њ—А–Є –Ї–Њ–љ—В—А–Њ–ї—М–љ–Њ–є –і–Є–µ—В–µ –Є –і–Є–µ—В–µ, –Њ–±–Њ–≥–∞—Й–µ–љ–љ–Њ–є –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–∞–Љ–Є (–і–Є–µ—В–∞ PlsEtn). –•—А–Њ–Љ–∞—В–Њ–≥—А–∞–Љ–Љ–∞ –њ–Њ–Ї–∞–Ј—Л–≤–∞–µ—В, —З—В–Њ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л (PlsEtn –Є PlsCho) –і–µ–є—Б—В–≤–Є—В–µ–ї—М–љ–Њ –њ—А–Є—Б—Г—В—Б—В–≤—Г—О—В –≤ –і–Є–µ—В–µ PlsEtn. –°–Њ–Ї—А–∞—Й–µ–љ–Є—П: —Б–Љ. —А–Є—Б. 1.
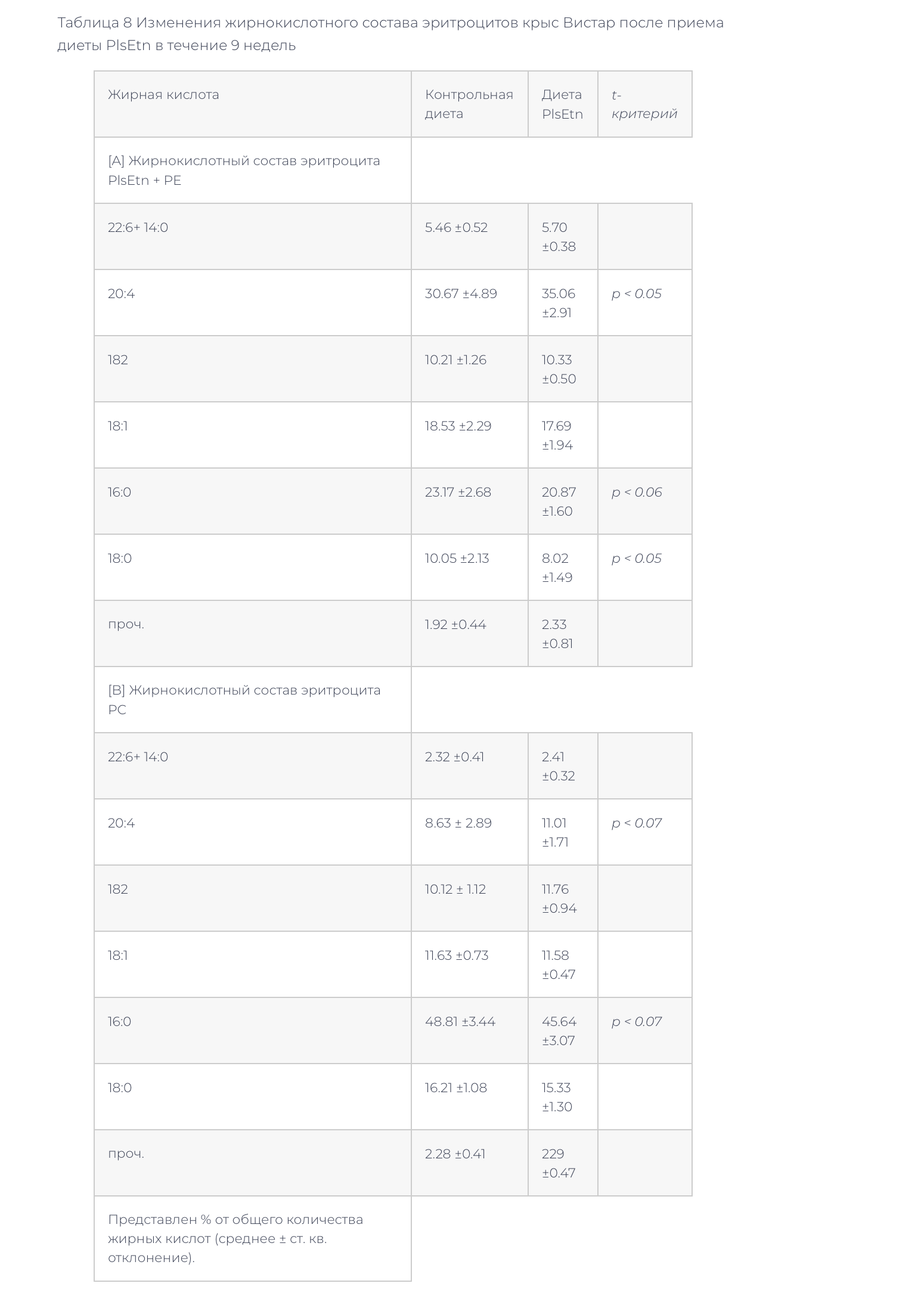
–Ф–Є–µ—В–∞ PlsEtn, –Ї–Њ—В–Њ—А—Г—О –Ї—А—Л—Б—Л ZDF –њ–Њ–ї—Г—З–∞–ї–Є –≤ —В–µ—З–µ–љ–Є–µ 4 –љ–µ–і–µ–ї—М, –Є–љ–і—Г—Ж–Є—А–Њ–≤–∞–ї–∞ –њ–Њ–≤—Л—И–µ–љ–Є–µ —Г—А–Њ–≤–љ—П PlsEtn –Є PE –Є —Б–љ–Є–ґ–µ–љ–Є–µ —Г—А–Њ–≤–љ—П PC –≤ –Љ–µ–Љ–±—А–∞–љ–∞—Е —Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤ (—В–∞–±–ї–Є—Ж–∞ 4). –Ф–Є–µ—В–∞ PlsEtn, –Ї–Њ—В–Њ—А—Г—О –Ї—А—Л—Б—Л –Т–Є—Б—В–∞—А –њ–Њ–ї—Г—З–∞–ї–Є –≤ —В–µ—З–µ–љ–Є–µ 9 –љ–µ–і–µ–ї—М, —В–∞–Ї–ґ–µ –Є–љ–і—Г—Ж–Є—А–Њ–≤–∞–ї–∞ –њ–Њ–≤—Л—И–µ–љ–Є–µ —Г—А–Њ–≤–љ—П PlsEtn –Є —В–µ–љ–і–µ–љ—Ж–Є—О –Ї —Б–љ–Є–ґ–µ–љ–Є—О PC –≤ –Љ–µ–Љ–±—А–∞–љ–∞—Е —Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤ (—В–∞–±–ї–Є—Ж–∞ 7). –Т –ґ–Є—А–љ–Њ–Ї–Є—Б–ї–Њ—В–љ–Њ–Љ —Б–Њ—Б—В–∞–≤–µ PlsEtn + PE –љ–∞–±–ї—О–і–∞–ї–Њ—Б—М —Г–≤–µ–ї–Є—З–µ–љ–Є–µ —Б–Њ–і–µ—А–ґ–∞–љ–Є—П ARA (20:4) –Є —Г–Љ–µ–љ—М—И–µ–љ–Є–µ —Б–Њ–і–µ—А–ґ–∞–љ–Є—П –Њ–ї–µ–Є–љ–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л (18:1) (—В–∞–±–ї–Є—Ж–∞ 8). –£–≤–µ–ї–Є—З–µ–љ–Є–µ —Б–Њ–і–µ—А–ґ–∞–љ–Є—П ARA –≤ PlsEtn –Љ–µ–Љ–±—А–∞–љ—Л —Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤ –Љ–Њ–ґ–µ—В –Њ—В—А–∞–ґ–∞—В—М –ґ–Є—А–љ–Њ–Ї–Є—Б–ї–Њ—В–љ—Л–є —Б–Њ—Б—В–∞–≤ PlsEtn –≤ –і–Є–µ—В–µ (—В–∞–±–ї–Є—Ж–∞ 1). –≠—В–Є —А–µ–Ј—Г–ї—М—В–∞—В—Л –∞–љ–∞–ї–Є–Ј–∞ –Љ–µ–Љ–±—А–∞–љ —Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤ —Б–≤–Є–і–µ—В–µ–ї—М—Б—В–≤—Г—О –Њ —В–Њ–Љ, —З—В–Њ –њ–Є—Й–µ–≤—Л–µ PlsEtn –Љ–Њ–≥—Г—В —Г–≤–µ–ї–Є—З–Є–≤–∞—В—М –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ—Г—О –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є—О PlsEtn –≤ —В–Ї–∞–љ—П—Е.
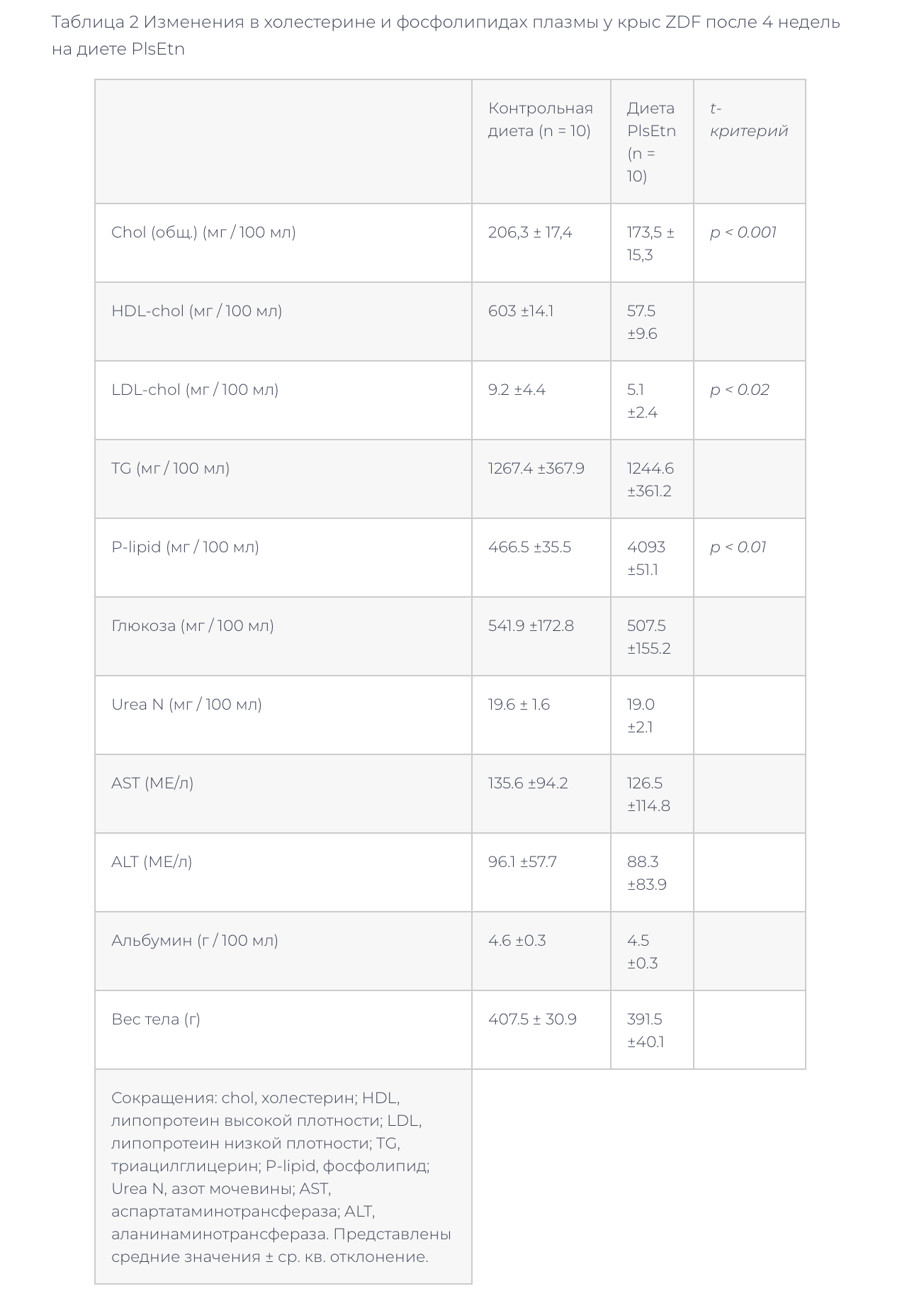
–Ґ–∞–±–ї–Є—Ж–∞ 2 –Ш–Ј–Љ–µ–љ–µ–љ–Є—П –≤ —Е–Њ–ї–µ—Б—В–µ—А–Є–љ–µ –Є —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–∞—Е –њ–ї–∞–Ј–Љ—Л —Г –Ї—А—Л—Б ZDF –њ–Њ—Б–ї–µ 4 –љ–µ–і–µ–ї—М –љ–∞ –і–Є–µ—В–µ PlsEtn
–•—А–Њ–Љ–∞—В–Њ–≥—А–∞–Љ–Љ–∞ —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–љ–Њ–≥–Њ –∞–љ–∞–ї–Є–Ј–∞ –і–Є–µ—В—Л PlsEtn –Є–Љ–µ–µ—В –±–Њ–ї—М—И–Њ–є –њ–Є–Ї PlsEtn –њ–Њ —Б—А–∞–≤–љ–µ–љ–Є—О —Б –Ї–Њ–љ—В—А–Њ–ї—М–љ–Њ–є –і–Є–µ—В–Њ–є. –≠—В–Њ –Њ–Ј–љ–∞—З–∞–µ—В, —З—В–Њ PlsEtn –і–µ–є—Б—В–≤–Є—В–µ–ї—М–љ–Њ —Б–Њ–і–µ—А–ґ–Є—В—Б—П –≤ –і–Є–µ—В–µ PlsEtn (—А–Є—Б. 2).
–Ф–Є–µ—В–∞, —Б–Њ–і–µ—А–ґ–∞–≤—И–∞—П 0,1% –Љ–∞—Б—Б. –Њ—З–Є—Й–µ–љ–љ—Л—Е –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ (–і–Є–µ—В–∞ PlsEtn), –≤—Л–Ј–≤–∞–ї–∞ —Г –Ї—А—Л—Б ZDF —Б–љ–Є–ґ–µ–љ–Є–µ —Г—А–Њ–≤–љ—П —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤ –Є —Е–Њ–ї–µ—Б—В–µ—А–Є–љ–∞ –≤ –њ–ї–∞–Ј–Љ–µ –≤ —В–µ—З–µ–љ–Є–µ 4 –љ–µ–і–µ–ї—М (—В–∞–±–ї–Є—Ж–∞ 2).
–Ю–і–љ–∞–Ї–Њ —А–µ–Ј—Г–ї—М—В–∞—В—Л –і—А—Г–≥–Є—Е —Б—В–∞–љ–і–∞—А—В–љ—Л—Е –ї–∞–±–Њ—А–∞—В–Њ—А–љ—Л—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є –њ–ї–∞–Ј–Љ—Л, –≤ —В–Њ–Љ —З–Є—Б–ї–µ –љ–∞ —В—А–Є–≥–ї–Є—Ж–µ—А–Њ–ї (TG), –≥–ї—О–Ї–Њ–Ј—Г, –∞–ї–∞–љ–Є–љ–∞–Љ–Є–љ–Њ—В—А–∞–љ—Б—Д–µ—А–∞–Ј—Г (ALT), –∞—Б–њ–∞—А—В–∞—В–∞–Љ–Є–љ–Њ—В—А–∞–љ—Б—Д–µ—А–∞–Ј—Г (AST), –∞–Ј–Њ—В –Љ–Њ—З–µ–≤–Є–љ—Л –Є –∞–ї—М–±—Г–Љ–Є–љ, –љ–µ –Њ—В–ї–Є—З–∞–ї–Є—Б—М (—В–∞–±–ї–Є—Ж–∞ 2). –Ь–µ–ґ–і—Г –≥—А—Г–њ–њ–∞–Љ–Є –љ–∞ —А–∞–Ј–љ–Њ–є –і–Є–µ—В–µ —В–∞–Ї–ґ–µ –љ–µ –±—Л–ї–Њ –Њ—В–Љ–µ—З–µ–љ–Њ —А–∞–Ј–ї–Є—З–Є–є –≤ –≤–µ—Б–µ —В–µ–ї–∞. –§–Њ—Б—Д–Њ–ї–Є–њ–Є–і–љ—Л–є —Б–Њ—Б—В–∞–≤ –њ–ї–∞–Ј–Љ—Л, –Є–Ј—Г—З–µ–љ–љ—Л–є –њ–Њ –љ–∞—И–µ–Љ—Г –Љ–µ—В–Њ–і—Г –Т–≠–Ц–•, –Є–Љ–µ–ї –њ–Њ–≤—Л—И–µ–љ–љ–Њ–µ —Б–Њ–і–µ—А–ґ–∞–љ–Є–µ SM –≤ –≥—А—Г–њ–њ–µ, –њ–Њ—Г—З–∞–≤—И–µ–є –і–Є–µ—В—Г PlsEtn (—В–∞–±–ї–Є—Ж–∞ 3).
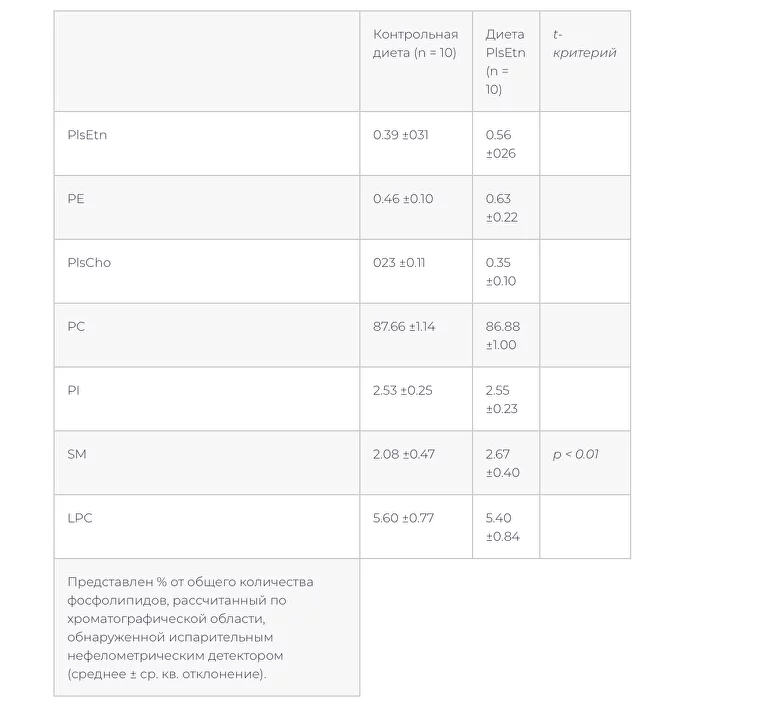
–Я–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л –≤ –њ–ї–∞–Ј–Љ–µ –±—Л–ї–Є –Њ–±–љ–∞—А—Г–ґ–µ–љ—Л —Б –њ–Њ–Љ–Њ—Й—М—О –љ–∞—И–µ–≥–Њ –Љ–µ—В–Њ–і–∞ –Т–≠–Ц–•, –љ–Њ –±—Л–ї–Є —Б–ї–Є—И–Ї–Њ–Љ –Љ–∞–ї—Л, —З—В–Њ–±—Л –Њ—Ж–µ–љ–Є—В—М –Є–Ј–Љ–µ–љ–µ–љ–Є—П –≤ –љ–Є—Е (—В–∞–±–ї–Є—Ж–∞ 3, —А–Є—Б. 3). –Ф–Є–µ—В–∞ PlsEtn –Є–љ–і—Г—Ж–Є—А–Њ–≤–∞–ї–∞ –њ–Њ–≤—Л—И–µ–љ–Є–µ —Г—А–Њ–≤–љ—П PlsEtn –Є PE —Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤ –Є —Б–љ–Є–ґ–µ–љ–Є–µ —Г—А–Њ–≤–љ—П PC —Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤ (—В–∞–±–ї–Є—Ж–∞ 4).
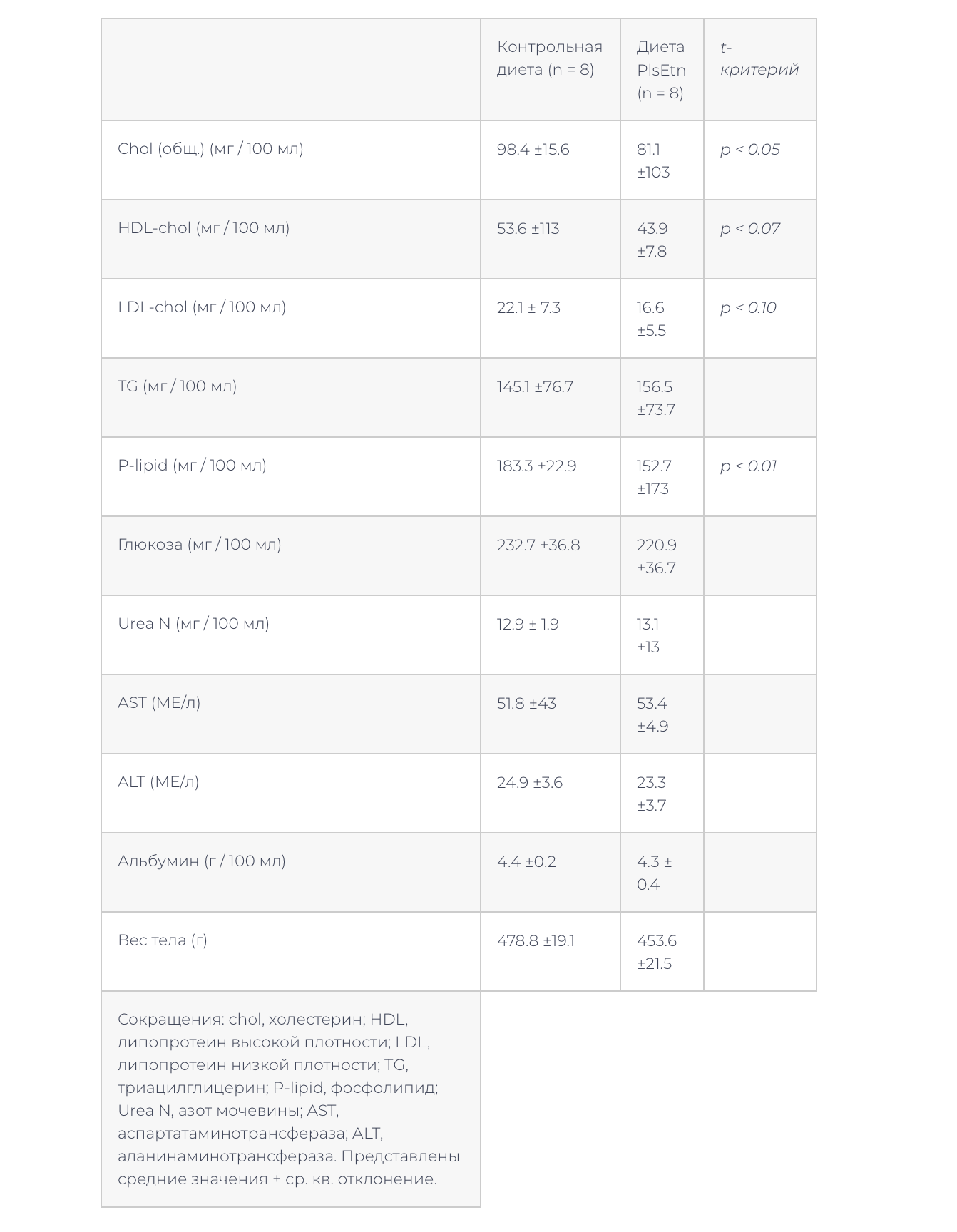
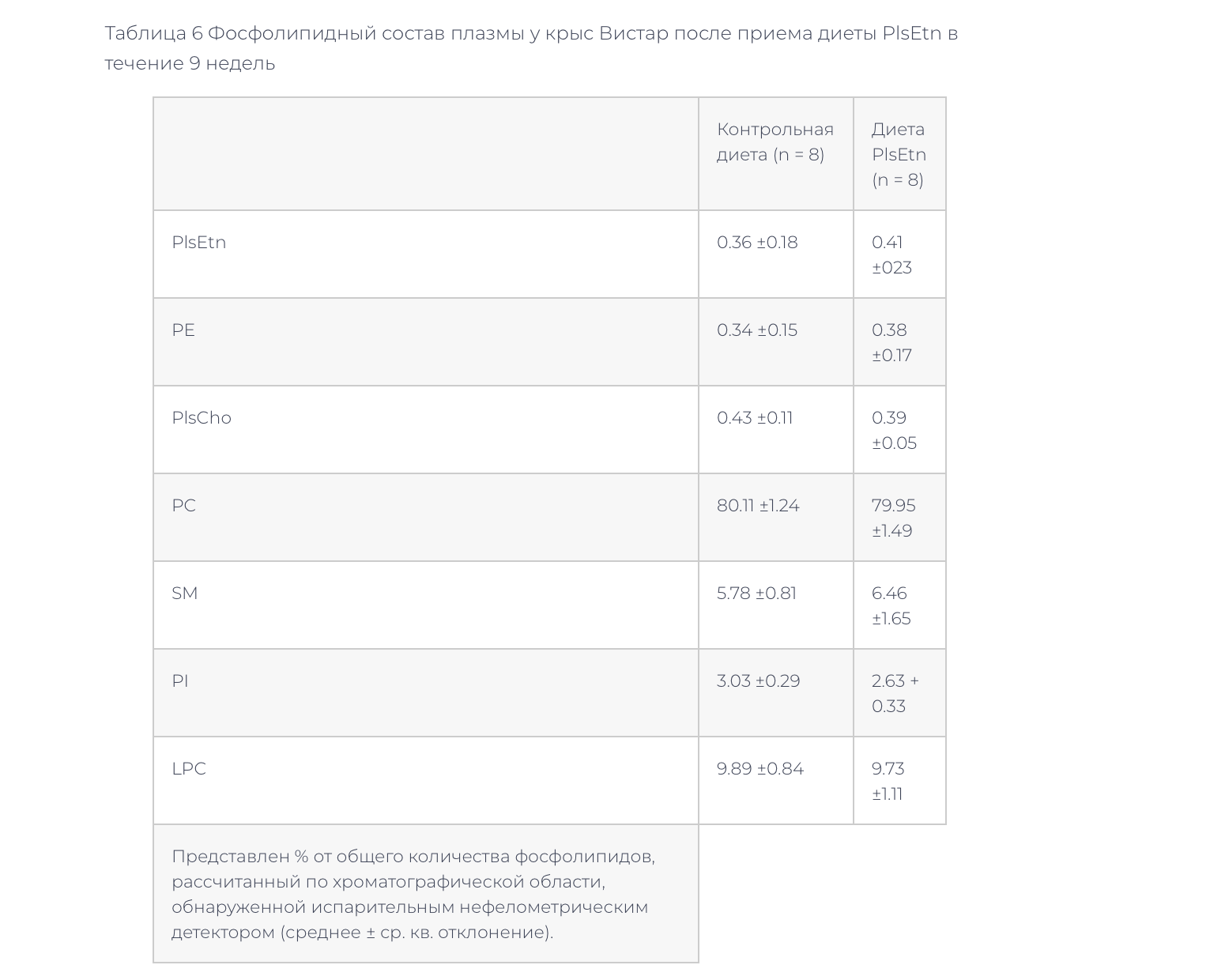
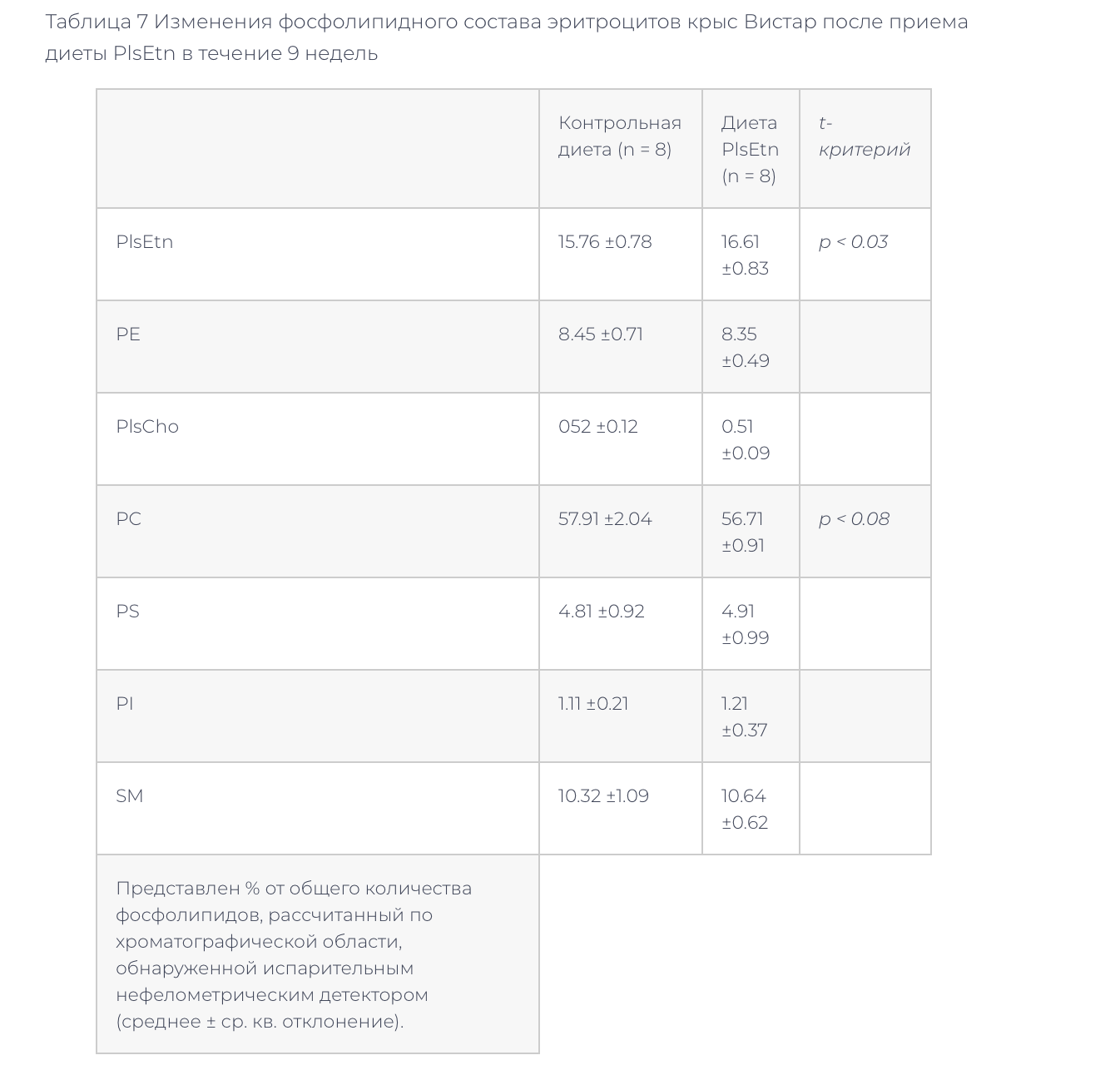
–Ч–і–Њ—А–Њ–≤—Л–µ –Ї—А—Л—Б—Л –ї–Є–љ–Є–Є –Т–Є—Б—В–∞—А –±—Л–ї–Є –њ–µ—А–µ–≤–µ–і–µ–љ—Л –љ–∞ —В—Г –ґ–µ –і–Є–µ—В—Г PlsEtn –љ–∞ 9 –љ–µ–і–µ–ї—М. –Ф–Є–µ—В–∞ PlsEtn –≤–љ–Њ–≤—М –њ—А–Є–≤–µ–ї–∞ –Ї —Б–љ–Є–ґ–µ–љ–Є—О —Г—А–Њ–≤–љ—П —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤ –Є —Е–Њ–ї–µ—Б—В–µ—А–Є–љ–∞ –≤ –њ–ї–∞–Ј–Љ–µ, –Њ–і–љ–∞–Ї–Њ —А–µ–Ј—Г–ї—М—В–∞—В—Л –і—А—Г–≥–Є—Е —Б—В–∞–љ–і–∞—А—В–љ—Л—Е –ї–∞–±–Њ—А–∞—В–Њ—А–љ—Л—Е –∞–љ–∞–ї–Є–Ј–Њ–≤ –њ–ї–∞–Ј–Љ—Л –љ–µ –Є–Ј–Љ–µ–љ–Є–ї–Є—Б—М (—В–∞–±–ї–Є—Ж–∞ 5). –§–Њ—Б—Д–Њ–ї–Є–њ–Є–і–љ—Л–є —Б–Њ—Б—В–∞–≤ –њ–ї–∞–Ј–Љ—Л –љ–µ –Є–Ј–Љ–µ–љ–Є–ї—Б—П (–Ґ–∞–±–ї–Є—Ж–∞ 6). –Т —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–љ–Њ–Љ —Б–Њ—Б—В–∞–≤–µ –Љ–µ–Љ–±—А–∞–љ —Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤ –≤ –≥—А—Г–њ–њ–µ –љ–∞ –і–Є–µ—В–µ PlsEtn –Њ—В–Љ–µ—З–µ–љ–Њ —Г–≤–µ–ї–Є—З–µ–љ–Є–µ PlsEtn –Є —В–µ–љ–і–µ–љ—Ж–Є—П –Ї —Б–љ–Є–ґ–µ–љ–Є—О PC (—В–∞–±–ї–Є—Ж–∞ 7).
–≠—В–Є —А–µ–Ј—Г–ї—М—В–∞—В—Л –њ–Њ–Ї–∞–Ј—Л–≤–∞—О—В, —З—В–Њ –њ–Є—Й–µ–≤—Л–µ PlsEtn –Є–љ–і—Г—Ж–Є—А—Г—О—В —Б–љ–Є–ґ–µ–љ–Є–µ —Г—А–Њ–≤–љ—П —Е–Њ–ї–µ—Б—В–µ—А–Є–љ–∞ –Є —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤ –≤ –њ–ї–∞–Ј–Љ–µ –Ї–∞–Ї –Ј–і–Њ—А–Њ–≤—Л—Е –Ї—А—Л—Б, —В–∞–Ї –Є –Ї—А—Л—Б ZDF. –Ш–Ј–≤–µ—Б—В–љ–Њ, —З—В–Њ –њ–Є—Й–µ–≤—Л–µ —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і—Л –≤—Л–Ј—Л–≤–∞—О—В —Б–љ–Є–ґ–µ–љ–Є–µ —Г—А–Њ–≤–љ—П —Е–Њ–ї–µ—Б—В–µ—А–Є–љ–∞ –≤ –њ–ї–∞–Ј–Љ–µ, –≤–µ—А–Њ—П—В–љ–Њ, –≤—Б–ї–µ–і—Б—В–≤–Є–µ –Є–љ–≥–Є–±–Є—А–Њ–≤–∞–љ–Є—П –Ї–Є—И–µ—З–љ–Њ–є –∞–±—Б–Њ—А–±—Ж–Є–Є —Е–Њ–ї–µ—Б—В–µ—А–Є–љ–∞ [6,9,10]. –°—Г—Й–µ—Б—В–≤—Г–µ—В –Є–љ—Д–Њ—А–Љ–∞—Ж–Є—П –Њ —В–Њ–Љ, —З—В–Њ –њ–Є—Й–µ–≤—Л–µ PE, –љ–Њ –љ–µ –њ–Є—Й–µ–≤—Л–µ PC, –≤—Л–Ј—Л–≤–∞–ї–Є —Б–љ–Є–ґ–µ–љ–Є–µ —Г—А–Њ–≤–љ—П —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤, —Е–Њ–ї–µ—Б—В–µ—А–Є–љ–∞ –Є –∞–њ–Њ–ї–Є–њ–Њ–њ—А–Њ—В–µ–Є–љ–∞ A-1 –≤ –њ–ї–∞–Ј–Љ–µ –Ї—А–Њ–≤–Є –Ї—А—Л—Б [11]. –Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, —Н—В–∞–љ–Њ–ї–∞–Љ–Є–љ-—Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і—Л, –љ–µ —В–Њ–ї—М–Ї–Њ PlsEtn, –≤—Л–Ј—Л–≤–∞—О—В —Б–љ–Є–ґ–µ–љ–Є—П —Г—А–Њ–≤–љ—П —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤ –≤ –њ–ї–∞–Ј–Љ–µ. –§–Њ—Б—Д–Њ–ї–Є–њ–Є–і–љ—Л–є —Б–Њ—Б—В–∞–≤ –њ–ї–∞–Ј–Љ—Л –Ј–і–Њ—А–Њ–≤—Л—Е –Ї—А—Л—Б (–Т–Є—Б—В–∞—А) –љ–µ –Є–Ј–Љ–µ–љ–Є–ї—Б—П –њ–Њ—Б–ї–µ 9 –љ–µ–і–µ–ї—М –љ–∞ –і–Є–µ—В–µ PlsEtn (—В–∞–±–ї–Є—Ж–∞ 6).
–Ф—А—Г–≥–Є–µ —Б—В–∞–љ–і–∞—А—В–љ—Л–µ –ї–∞–±–Њ—А–∞—В–Њ—А–љ—Л–µ –∞–љ–∞–ї–Є–Ј—Л –њ–ї–∞–Ј–Љ—Л —В–∞–Ї–ґ–µ –Њ—Б—В–∞–ї–Є—Б—М –±–µ–Ј –Є–Ј–Љ–µ–љ–µ–љ–Є–є (—В–∞–±–ї–Є—Ж–∞ 5). –Т–µ—Б —В–µ–ї–∞ —В–∞–Ї–ґ–µ –љ–µ –Є–Ј–Љ–µ–љ–Є–ї—Б—П –њ–Њ—Б–ї–µ 9 –љ–µ–і–µ–ї—М –љ–∞ –і–Є–µ—В–µ PlsEtn (—В–∞–±–ї–Є—Ж–∞ 5).
–≠—В–Є —А–µ–Ј—Г–ї—М—В–∞—В—Л –њ–Њ–Ї–∞–Ј—Л–≤–∞—О—В, —З—В–Њ –њ–Є—Й–µ–≤—Л–µ PlsEtn –љ–µ –Њ–Ї–∞–Ј–∞–ї–Є –≤—А–µ–і–љ–Њ–≥–Њ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є—П –і–ї—П –Ј–і–Њ—А–Њ–≤—Л—Е –Ї—А—Л—Б –љ–µ—Б–Љ–Њ—В—А—П –љ–∞ —Б–љ–Є–ґ–µ–љ–Є–µ —Г—А–Њ–≤–љ—П —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤ –Є —Е–Њ–ї–µ—Б—В–µ—А–Є–љ–∞ –≤ –њ–ї–∞–Ј–Љ–µ. –Ш–љ—В–µ—А–µ—Б–љ–Њ –Њ—В–Љ–µ—В–Є—В—М, —З—В–Њ —Г—А–Њ–≤–µ–љ—М —В—А–Є–∞—Ж–Є–ї–≥–ї–Є—Ж–µ—А–Њ–ї–∞ –њ–ї–∞–Ј–Љ—Л –љ–µ –Є–Ј–Љ–µ–љ–Є–ї—Б—П —Г –Ї—А—Л—Б ZDF –Є –Ї—А—Л—Б –Т–Є—Б—В—А–µ –≤—Б–ї–µ–і—Б—В–≤–Є–µ –њ—А–Є–µ–Љ–∞ –і–Є–µ—В—Л PlsEtn –љ–µ—Б–Љ–Њ—В—А—П –љ–∞ —Б–љ–Є–ґ–µ–љ–Є–µ —Г—А–Њ–≤–љ—П —Е–Њ–ї–µ—Б—В–µ—А–Є–љ–∞ –Є —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤.
–С–Њ–ї—М—И–Є–љ—Б—В–≤–Њ –і–Є–µ—В–Є—З–µ—Б–Ї–Є—Е –≥–ї–Є—Ж–µ—А–Њ—Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤ –Љ–Њ–≥—Г—В –±—Л—В—М –≥–Є–і—А–Њ–ї–Є–Ј–Њ–≤–∞–љ—Л –≤ –њ–Њ–ї–Њ–ґ–µ–љ–Є–Є SN-2 –њ–∞–љ–Ї—А–µ–∞—В–Є—З–µ—Б–Ї–Њ–є —Д–Њ—Б—Д–Њ–ї–Є–њ–∞–Ј–Њ–є –Р2 –≤ –њ—А–Њ—Б–≤–µ—В–µ –Ї–Є—И–µ—З–љ–Є–Ї–∞, –∞ –Ј–∞—В–µ–Љ –∞–±—Б–Њ—А–±–Є—А–Њ–≤–∞–љ—Л —Н–љ—В–µ—А–Њ—Ж–Є—В–∞–Љ–Є –≤ –Ї–∞—З–µ—Б—В–≤–µ —Б–≤–Њ–±–Њ–і–љ—Л—Е –ґ–Є—А–љ—Л—Е –Ї–Є—Б–ї–Њ—В –Є –ї–Є–Ј–Њ—Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤ [6,12].
–°—Г—Й–µ—Б—В–≤—Г–µ—В –Є–љ—Д–Њ—А–Љ–∞—Ж–Є—П –Њ –њ—А–Є—Б—Г—В—Б—В–≤–Є–Є –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ-–∞–Ї—В–Є–≤–љ–Њ–є —Д–Њ—Б—Д–Њ–ї–Є–њ–∞–Ј—Л –Р2 –≤ —Н–њ–Є—В–µ–ї–Є–Є —В–Њ–љ–Ї–Њ–є –Ї–Є—И–Ї–Є [13]. –Я–Њ—Б–Ї–Њ–ї—М–Ї—Г –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л —Б–Њ–і–µ—А–ґ–∞—В –≤–Є–љ–Є–ї—Н—Д–Є—А–љ—Г—О —Б–≤—П–Ј—М –≤ –њ–Њ–ї–Њ–ґ–µ–љ–Є–Є sn-1 –≥–ї–Є—Ж–µ—А–Є–љ–∞, –ї–Є–Ј–Њ—Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і—Л, –њ–Њ–ї—Г—З–µ–љ–љ—Л–µ –Є–Ј –њ–Є—Й–µ–≤—Л—Е –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ –њ–Њ—Б–ї–µ –њ–µ—А–µ–≤–∞—А–Є–≤–∞–љ–Є—П —Д–Њ—Б—Д–Њ–ї–Є–њ–∞–Ј–Њ–є –Р2, —Б–Њ–і–µ—А–ґ–∞—В –≤–Є–љ–Є–ї—Н—Д–Є—А–љ—Г—О —Б–≤—П–Ј—М. –Ш–Ј–≤–µ—Б—В–љ–Њ, —З—В–Њ –њ–Є—Й–µ–≤—Л–µ –∞–ї–Ї–Є–ї—М–љ—Л–µ –≥–ї–Є—Ж–µ—А–Њ–ї—Л –≤—Б–∞—Б—Л–≤–∞—О—В—Б—П –Є–Ј –њ–Є—Й–µ–≤–∞—А–Є—В–µ–ї—М–љ–Њ–≥–Њ —В—А–∞–Ї—В–∞ –љ–µ–Є–Ј–Љ–µ–љ–љ—Л–Љ–Є –±–µ–Ј —А–∞—Б—Й–µ–њ–ї–µ–љ–Є—П —Н—Д–Є—А–љ–Њ–є —Б–≤—П–Ј–Є [14-18].
–Ю–і–љ–∞–Ї–Њ –љ–µ–Є–Ј–≤–µ—Б—В–љ–Њ, –≤—Б–∞—Б—Л–≤–∞–µ—В—Б—П –ї–Є –љ–µ–њ–Њ–≤—А–µ–ґ–і–µ–љ–љ–Њ–є –≤–Є–љ–Є–ї—Н—Д–Є—А–љ–∞—П —Б–≤—П–Ј—М –њ–Є—Й–µ–≤—Л—Е —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤. –Т–Є–љ–Є–ї—Н—Д–Є—А–љ–∞—П —Б–≤—П–Ј—М (1-0 –∞–ї–Ї-1вА≤-—Н–љ–Є–ї-sn-–≥–ї–Є—Ж–µ—А–Њ–ї) –љ–µ —В–Њ–ґ–і–µ—Б—В–≤–µ–љ–љ–∞ —Н—Д–Є—А–љ–Њ–є —Б–≤—П–Ј–Є –∞–ї–Ї–Є–ї—М–љ—Л—Е –≥–ї–Є—Ж–µ—А–Њ–ї–Њ–≤ (1-0 –∞–ї–Ї–Є–ї-sn-–≥–ї–Є—Ж–µ—А–Њ–ї). –Ш–Ј–≤–µ—Б—В–љ–Њ, —З—В–Њ –≤–Є–љ–Є–ї—Н—Д–Є—А–љ–∞—П —Б–≤—П–Ј—М –Њ—Б–Њ–±–µ–љ–љ–Њ —З—Г–≤—Б—В–≤–Є—В–µ–ї—М–љ–∞ –Ї HCl [7,8]. –°–Њ–Њ–±—Й–∞–µ—В—Б—П –Њ–± –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–Є, –≤ –Ї–Њ—В–Њ—А–Њ–Љ –њ—А–Є –і–Є–µ—В–µ, —Б–Њ–і–µ—А–ґ–∞—Й–µ–є 10% —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤ –Љ–Њ–Ј–≥–∞ –±—Л–Ї–∞, –љ–µ –љ–∞–±–ї—О–і–∞–ї–Њ—Б—М –і–µ–≥—А–∞–і–∞—Ж–Є–Є –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ –≤ –ї–∞–±–Њ—А–∞—В–Њ—А–љ—Л—Е —Г—Б–ї–Њ–≤–Є—П—Е, –Є–Љ–Є—В–Є—А—Г—О—Й–Є—Е –ґ–µ–ї—Г–і–Њ–Ї –Є –њ—А–Њ—Б–≤–µ—В —В–Њ–љ–Ї–Њ–≥–Њ –Ї–Є—И–µ—З–љ–Є–Ї–∞, –∞ –њ—А–Є–µ–Љ 10% —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤ –Љ–Њ–Ј–≥–∞ –±—Л–Ї–∞ –њ—А–Є–≤–µ–ї –Ї –њ–Њ–≤—Л—И–µ–љ–Є—О —Г—А–Њ–≤–љ—П –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ –≤ –њ–ї–∞–Ј–Љ–µ –Ї—А–Њ–≤–Є [19].
–Я–Њ—Н—В–Њ–Љ—Г –≤–µ—А–Њ—П—В–љ–Њ, —З—В–Њ –ї–Є–Ј–Њ—Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і—Л (–ї–Є–Ј–Њ–њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л), —Б–Њ–і–µ—А–ґ–∞—Й–Є–µ –≤–Є–љ–Є–ї—Н—Д–Є—А–љ—Г—О —Б–≤—П–Ј—М, –њ–Њ–ї—Г—З–µ–љ–љ—Г—О –Є–Ј –њ–Є—Й–µ–≤–Њ–≥–Њ PlsEtn, –≤—Б–∞—Б—Л–≤–∞—О—В—Б—П –љ–µ–Є–Ј—Б–µ–љ–љ—Л–Љ–Є –Є–Ј –Ї–Є—И–µ—З–љ–Є–Ї–∞ –Ї—А—Л—Б—Л.
–Ґ–∞–±–ї–Є—Ж–∞ 3 –§–Њ—Б—Д–Њ–ї–Є–њ–Є–і–љ—Л–є —Б–Њ—Б—В–∞–≤ –њ–ї–∞–Ј–Љ—Л —Г –Ї—А—Л—Б ZDF –њ–Њ—Б–ї–µ –њ—А–Є–µ–Љ–∞ –і–Є–µ—В—Л PIsEtn –≤ —В–µ—З–µ–љ–Є–µ 4 –љ–µ–і–µ–ї—М
–Я–Њ—Б–Ї–Њ–ї—М–Ї—Г 1-0-–∞–ї–Ї–Є–ї–≥–ї–Є—Ж–µ—А–Њ–ї –њ—А–Є–љ–Є–Љ–∞–µ—В —Г—З–∞—Б—В–Є–µ –≤ –±–Є–Њ—Б–Є–љ—В–µ–Ј–µ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ –њ–Њ—Б–ї–µ –њ–µ—А–Њ–Ї—Б–Є—Б–Њ–Љ–∞–ї—М–љ–Њ–≥–Њ —Н—В–∞–њ–∞, —Б–Њ–Њ–±—Й–∞–µ—В—Б—П –Њ –ї–µ—З–µ–љ–Є–Є –љ–∞—Б–ї–µ–і—Б—В–≤–µ–љ–љ—Л—Е –њ–µ—А–Њ–Ї—Б–Є—Б–Њ–Љ–љ—Л—Е –њ–∞—В–Њ–ї–Њ–≥–Є–є, –Њ—Б–љ–Њ–≤–∞–љ–љ–Њ–Љ –љ–∞ –Ј–∞–Љ–µ—Й–µ–љ–Є–Є –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤, —Б –њ–Њ–Љ–Њ—Й—М—О –њ–Є—Й–µ–≤—Л—Е 1-0 –∞–ї–Ї–Є–ї–≥–ї–Є—Ж–µ—А–Њ–ї–Њ–≤, –Ї–Њ—В–Њ—А–Њ–µ –њ—А–Є–≤–µ–ї–Њ –Ї –њ–Њ–≤—Л—И–µ–љ–Є—О —Г—А–Њ–≤–љ—П –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ –≤ –љ–µ–Ї–Њ—В–Њ—А—Л—Е —В–Ї–∞–љ—П—Е [15-18,20] ,
–Т —А–∞–±–Њ—В–µ (Wood et al.) [20,21] –Њ–њ–Є—Б—Л–≤–∞–µ—В—Б—П —Г—Б–њ–µ—И–љ—Л–є —Б–Є–љ—В–µ–Ј –њ—А–µ–і—И–µ—Б—В–≤–µ–љ–љ–Є–Ї–∞ –∞–ї–Ї–Є–ї-–і–Є–∞—Ж–Є–ї—М–љ–Њ–≥–Њ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–∞ —Б –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–µ–Љ –њ–∞–ї—М–Љ–Є—В–Є–љ–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л –≤ sn-1, DHA –≤ sn-2 –Є –ї–Є–њ–Њ–µ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л –≤ sn-3 (PPI-1011).
–°–Њ–Њ–±—Й–∞–µ—В—Б—П, —З—В–Њ PPI-1011 –њ–Њ–њ–Њ–ї–љ–Є–ї PlsEtn –Ї—Г–ї—М—В–Є–≤–Є—А—Г–µ–Љ—Л—Е –ї–Є–Љ—Д–Њ—Ж–Є—В–Њ–≤ –њ–∞—Ж–Є–µ–љ—В–Њ–≤ c RCDP [20], –∞ –њ–µ—А–Њ—А–∞–ї—М–љ–Њ–µ –≤–≤–µ–і–µ–љ–Є–µ PPI-1011 –Ї—А–Њ–ї–Є–Ї–∞–Љ –≤ —В–µ—З–µ–љ–Є–µ 2 –љ–µ–і–µ–ї—М –њ–Њ–≤—Л—Б–Є–ї–Њ —Г—А–Њ–≤–µ–љ—М DHA –Є DHA-—Б–Њ–і–µ—А–ґ–∞—Й–Є—Е PlsEtn –≤ –њ–ї–∞–Ј–Љ–µ –Є —Б–µ—В—З–∞—В–Ї–µ –≥–ї–∞–Ј–∞ [21].
–Я—А–Є–µ–Љ –Ї—А—Л—Б–∞–Љ–Є –і–Є–µ—В—Л PlsEtn —Г–≤–µ–ї–Є—З–Є–ї –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ–µ —Б–Њ–і–µ—А–ґ–∞–љ–Є–µ PlsEtn –≤ –Љ–µ–Љ–±—А–∞–љ–∞—Е —Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤ –≤ —В–µ—З–µ–љ–Є–µ 4 –љ–µ–і–µ–ї—М, –∞ –њ—А–Є–µ–Љ —В–Њ–є –ґ–µ –і–Є–µ—В—Л –Ј–і–Њ—А–Њ–≤—Л–Љ–Є –Ї—А—Л—Б–∞–Љ–Є –≤ —В–µ—З–µ–љ–Є–µ 9 –љ–µ–і–µ–ї—М —В–∞–Ї–ґ–µ —Г–≤–µ–ї–Є—З–Є–ї –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ–µ —Б–Њ–і–µ—А–ґ–∞–љ–Є–µ PlsEtn –≤ —Н—А–Є—В—А–Њ—Ж–Є—В–∞—Е.
–Э–∞—Б—В–Њ—П—Й–µ–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–µ –њ–Њ–Ї–∞–Ј—Л–≤–∞–µ—В, —З—В–Њ –њ–Є—Й–µ–≤–Њ–є PlsEtn –±–µ–Ј–Њ–њ–∞—Б–µ–љ –і–ї—П –Ј–і–Њ—А–Њ–≤—Л—Е –Ї—А—Л—Б, –љ–µ—Б–Љ–Њ—В—А—П –љ–∞ —Б–љ–Є–ґ–µ–љ–Є–µ —Г—А–Њ–≤–љ—П —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤ –њ–ї–∞–Ј–Љ—Л. –Я–µ—А–Њ—А–∞–ї—М–љ–Њ–µ –≤–≤–µ–і–µ–љ–Є–µ –Њ—З–Є—Й–µ–љ–љ—Л—Е –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ –Љ–Њ–ґ–µ—В –±—Л—В—М –±–Њ–ї–µ–µ —Н—Д—Д–µ–Ї—В–Є–≤–љ—Л–Љ –њ—А–Є –њ–Њ–њ–Њ–ї–љ–µ–љ–Є–Є –Є/–Є–ї–Є –Ј–∞–Љ–µ—Й–µ–љ–Є–Є –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ —В–Ї–∞–љ–µ–є, —З–µ–Љ –њ–µ—А–Њ—А–∞–ї—М–љ–Њ–µ –≤–≤–µ–і–µ–љ–Є–µ –∞–ї–Ї–Є–ї—М–љ—Л—Е –≥–ї–Є—Ж–µ—А–Њ–ї–Њ–≤ –Є–ї–Є –њ—А–µ–і—И–µ—Б—В–≤–µ–љ–љ–Є–Ї–∞ –∞–ї–Ї–Є–ї-–і–Є–∞—Ж–Є–ї—М–љ—Л—Е –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ (PPI-1011), –њ–Њ—В–Њ–Љ—Г —З—В–Њ –≤–≤–Њ–і–Є–Љ—Л–є –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ –Љ–Њ–ґ–µ—В —Б—В–∞—В—М –Є—Б—В–Њ—З–љ–Є–Ї–Њ–Љ –≥–ї–Є—Ж–µ—А–Њ–ї–∞ —Б –≤–Є–љ–Є–ї—Н—Д–Є—А–љ—Л–Љ–Є —Б–≤—П–Ј—П–Љ–Є, —Н—В–∞–љ–Њ–ї–∞–Љ–Є–љ–∞, —Е–Њ–ї–Є–љ–∞ –Є –њ–Њ–ї–Є–љ–µ–љ–∞—Б—Л—Й–µ–љ–љ—Л—Е –ґ–Є—А–љ—Л—Е –Ї–Є—Б–ї–Њ—В, —В–∞–Ї–Є—Е –Ї–∞–Ї DHA –Є ARA, –Є –≤—Б–µ —Н—В–Є –≤–µ—Й–µ—Б—В–≤–∞ –Љ–Њ–≥—Г—В –±—Л—В—М –њ–Њ–≤—В–Њ—А–љ–Њ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ—Л –≤ –±–Є–Њ—Б–Є–љ—В–µ–Ј–µ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ –≤ —А–∞–Ј–ї–Є—З–љ—Л—Е —В–Ї–∞–љ—П—Е.
–Э–µ–і–∞–≤–љ–Њ –Љ—Л —Б–Њ–Њ–±—Й–∞–ї–Є, —З—В–Њ –њ—А–µ–і–≤–∞—А–Є—В–µ–ї—М–љ–Њ–µ –≤–љ—Г—В—А–Є–±—А—О—И–Є–љ–љ–Њ–µ –≤–≤–µ–і–µ–љ–Є–µ –Њ—З–Є—Й–µ–љ–љ–Њ–≥–Њ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–∞ –Є–Ј –Љ—Л—И—Ж—Л –Ї—Г—А–Є–љ–Њ–є –≥—А—Г–і–Ї–Є (—Б–Њ–і–µ—А–ґ–Є—В 47,6% PlsEtn –Є 49,3% PlsCho) –Њ—Б–ї–∞–±–ї—П–µ—В –Є–љ–і—Г—Ж–Є—А–Њ–≤–∞–љ–љ–Њ–µ –ї–Є–њ–Њ–њ–Њ–ї–Є—Б–∞—Е–∞—А–Є–і–Њ–Љ –љ–µ–є—А–Њ–≤–Њ—Б–њ–∞–ї–µ–љ–Є–µ [22].
–Ю–і–љ–∞–Ї–Њ –њ–Њ–Ї–∞ –љ–µ —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ–Њ, –њ–Њ–≤—Л—И–∞—О—В –ї–Є –њ–Є—Й–µ–≤—Л–µ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л —Б–Њ–і–µ—А–ґ–∞–љ–Є–µ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ –≤ —В–Ї–∞–љ—П—Е –Љ–Њ–Ј–≥–∞ –њ—А–Є –і–µ—Д–Є—Ж–Є—В–µ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤.
–Ь–∞—В–µ—А–Є–∞–ї—Л –Є –Љ–µ—В–Њ–і—Л
–Я–Њ–ї—Г—З–µ–љ–Є–µ –Њ—З–Є—Й–µ–љ–љ—Л—Е –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤
–Я–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л –±—Л–ї–Є –њ–Њ–ї—Г—З–µ–љ—Л –Є–Ј –Ї—Г—А–Є–љ–Њ–є –Ї–Њ–ґ–Є. –Ъ—Г—А–Є–љ–∞—П –Ї–Њ–ґ–∞ –±—Л–ї–∞ –њ—А–Є–Њ–±—А–µ—В–µ–љ–∞ –љ–∞ —А—Л–љ–Ї–µ. –С–Њ–ї—М—И–∞—П —З–∞—Б—В—М –ґ–Є—А–∞ –±—Л–ї–∞ —Б–њ–µ—А–≤–∞ —Г–і–∞–ї–µ–љ–∞ —Б –Ї–Њ–ґ–Є –≤ –њ–∞—А–Њ–≤–Њ–є –њ–µ—З–Є, –Ј–∞—В–µ–Љ –Ї–Њ–ґ–∞ –±—Л–ї–∞ –≤—Л—Б—Г—И–µ–љ–∞ –≤—Л–Љ–Њ—А–∞–ґ–Є–≤–∞–љ–Є–µ–Љ. –Я–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л –±—Л–ї–Є –њ–Њ–ї—Г—З–µ–љ—Л –Є–Ј —Б—Г–±–ї–Є–Љ–Є—А–Њ–≤–∞–љ–љ–Њ–є –Ї–Њ–ґ–Є –њ–Њ —А–∞–љ–µ–µ –Њ–њ–Є—Б–∞–љ–љ–Њ–є –Љ–µ—В–Њ–і–Є–Ї–µ [23].–†–Є—Б—Г–љ–Њ–Ї 3 –†–µ–њ—А–µ–Ј–µ–љ—В–∞—В–Є–≤–љ–∞—П –Т–≠–Ц–•-—Е—А–Њ–Љ–∞—В–Њ–≥—А–∞–Љ–Љ–∞ —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤ –њ–ї–∞–Ј–Љ—Л –Ї—А—Л—Б –Т–Є—Б—В–∞—А. –Я–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л (PlsEtn –Є PlsCho) –≤ –њ–ї–∞–Ј–Љ–µ –Ї—А—Л—Б –±—Л–ї–Є –Њ–±–љ–∞—А—Г–ґ–µ–љ—Л –Њ–њ–Є—Б–∞–љ–љ—Л–Љ –љ–∞–Љ–Є –Љ–µ—В–Њ–і–Њ–Љ (A), –љ–Њ –±—Л–ї–Є —Б–ї–Є—И–Ї–Њ–Љ –Љ–∞–ї—Л, —З—В–Њ–±—Л –Њ—Ж–µ–љ–Є—В—М –Є–Ј–Љ–µ–љ–µ–љ–Є—П –≤ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–∞—Е, –љ–µ—Б–Љ–Њ—В—А—П –љ–∞ —Г–≤–µ–ї–Є—З–µ–љ–Є–µ —Е—А–Њ–Љ–∞—В–Њ–≥—А–∞—Д–∞ (B). LPC, –ї–Є–Ј–Њ—Д–Њ—Б—Д–∞—В–Є–і–Є–ї—Е–Њ–ї–Є–љ; –і—А—Г–≥–Є–µ –∞–±–±—А–µ–≤–Є–∞—В—Г—А—Л вАУ —Б–Љ. —А–Є—Б. 1.
–Ц–Є–≤–Њ—В–љ—Л–µ –Є –і–Є–µ—В—Л
–Ф–Є–µ—В—Г, —Б–Њ–і–µ—А–ґ–∞—Й–∞—П 0,1% –Љ–∞—Б—Б. –Њ—З–Є—Й–µ–љ–љ—Л—Е –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ (–і–Є–µ—В–∞ PlsEtn), –њ–Њ–ї—Г—З–∞–ї–Є –њ—Г—В–µ–Љ –і–Њ–±–∞–≤–ї–µ–љ–Є—П –Њ—З–Є—Й–µ–љ–љ—Л—Е –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ –Ї –Ї–Њ–љ—В—А–Њ–ї—М–љ–Њ–є –і–Є–µ—В–µ (AIN 96). –Ф–Є–µ—В—Л –±—Л–ї–Є –њ—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ—Л –Є –њ–Њ—Б—В–∞–≤–ї–µ–љ—Л –Ї–Њ–Љ–њ–∞–љ–Є–µ–є Oriental Yeast Co (–Ґ–Њ–Ї–Є–Њ, –ѓ–њ–Њ–љ–Є—П).–Т—Б–µ —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В—Л, —Б–≤—П–Ј–∞–љ–љ—Л–µ —Б –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–µ–Љ –ґ–Є–≤–Њ—В–љ—Л—Е, –±—Л–ї–Є –њ—А–Њ–≤–µ–і–µ–љ—Л –≤ —Б–Њ–Њ—В–≤–µ—В—Б—В–≤–Є–Є —Б –†—Г–Ї–Њ–≤–Њ–і—П—Й–Є–Љ–Є –њ—А–Є–љ—Ж–Є–њ–∞–Љ–Є –њ–Њ —Г—Е–Њ–і—Г –Є –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—О –ґ–Є–≤–Њ—В–љ—Л—Е –§–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–≥–Њ –Њ–±—Й–µ—Б—В–≤–∞ –ѓ–њ–Њ–љ–Є–Є.
–С—Л–ї–Є –њ—А–Є–ї–Њ–ґ–µ–љ—Л –≤—Б–µ —Г—Б–Є–ї–Є—П –Ї —В–Њ–Љ—Г, —З—В–Њ–±—Л —Б–≤–µ—Б—В–Є –Ї –Љ–Є–љ–Є–Љ—Г–Љ—Г —Б—В—А–∞–і–∞–љ–Є—П –ґ–Є–≤–Њ—В–љ—Л—Е –Є –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ –ґ–Є–≤–Њ—В–љ—Л—Е, –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–љ—Л—Е –≤ —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞—Е.
–Т—Б–µ –Ї—А—Л—Б—Л –±—Л–ї–Є –њ—А–Є–Њ–±—А–µ—В–µ–љ—Л –≤ –Ї–Њ–Љ–њ–∞–љ–Є–Є KBT Oriental Co. (–Ґ–Њ—Б—Г, –њ—А–µ—Д–µ–Ї—В—Г—А–∞ –°–∞–≥–∞, –ѓ–њ–Њ–љ–Є—П). –Ф–≤–∞–і—Ж–∞—В—М —Б–∞–Љ—Ж–Њ–≤ –Ї—А—Л—Б—Л ZDF –≤ –≤–Њ–Ј—А–∞—Б—В–µ 4 –љ–µ–і–µ–ї—М –њ–Њ–ї—Г—З–∞–ї–Є –Ї–Њ–љ—В—А–Њ–ї—М–љ—Г—О –і–Є–µ—В—Г –≤ —В–µ—З–µ–љ–Є–µ 10 –і–љ–µ–є –Є –±—Л–ї–Є —А–∞–Ј–і–µ–ї–µ–љ—Л –љ–∞ –і–≤–µ –≥—А—Г–њ–њ—Л —Б–ї—Г—З–∞–є–љ—Л–Љ –Њ–±—А–∞–Ј–Њ–Љ.
–Ю–і–љ–∞ –≥—А—Г–њ–њ–∞ (10 –Ї—А—Л—Б) –њ–Њ–ї—Г—З–∞–ї–∞ –Ї–Њ–љ—В—А–Њ–ї—М–љ—Г—О –і–Є–µ—В—Г –≤ —В–µ—З–µ–љ–Є–µ –µ—Й–µ 4 –љ–µ–і–µ–ї—М (–≥—А—Г–њ–њ–∞ –љ–∞ –Ї–Њ–љ—В—А–Њ–ї—М–љ–Њ–є –і–Є–µ—В–µ), –∞ –Њ—Б—В–∞–ї—М–љ—Л–µ –≥—А—Г–њ–њ—Л (10 –Ї—А—Л—Б) –њ–Њ–ї—Г—З–∞–ї–Є –і–Є–µ—В—Г PlsEtn (–≥—А—Г–њ–њ–∞ –љ–∞ –і–Є–µ—В–µ PlsEtn) –≤ —В–µ—З–µ–љ–Є–µ 4 –љ–µ–і–µ–ї—М. –Т –і—А—Г–≥–Њ–Љ —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–µ 18 —Б–∞–Љ—Ж–Њ–≤ –Ї—А—Л—Б –ї–Є–љ–Є–Є –Т–Є—Б—В–∞—А –≤ –≤–Њ–Ј—А–∞—Б—В–µ 6 –љ–µ–і–µ–ї—М –њ–Њ–ї—Г—З–∞–ї–Є –Ї–Њ–љ—В—А–Њ–ї—М–љ—Г—О –і–Є–µ—В—Г –≤ —В–µ—З–µ–љ–Є–µ 10 –і–љ–µ–є –Є –±—Л–ї–Є —А–∞–Ј–і–µ–ї–µ–љ—Л –љ–∞ –і–≤–µ –≥—А—Г–њ–њ—Л.
–Ю–і–љ–∞ –≥—А—Г–њ–њ–∞ –њ–Њ–ї—Г—З–∞–ї–∞ –Ї–Њ–љ—В—А–Њ–ї—М–љ—Г—О –і–Є–µ—В—Г –≤ —В–µ—З–µ–љ–Є–µ –µ—Й–µ 9 –љ–µ–і–µ–ї—М (–≥—А—Г–њ–њ–∞ –љ–∞ –Ї–Њ–љ—В—А–Њ–ї—М–љ–Њ–є –і–Є–µ—В–µ), –∞ –і—А—Г–≥–∞—П –≥—А—Г–њ–њ–∞ –њ–Њ–ї—Г—З–∞–ї–∞ –і–Є–µ—В—Г PlsEtn –≤ —В–µ—З–µ–љ–Є–µ 9 –љ–µ–і–µ–ї—М. –Т—Б–µ –Ї—А—Л—Б—Л –±—Л–ї–Є —А–∞–Ј–Љ–µ—Й–µ–љ—Л –≤ –Ї–ї–µ—В–Ї–∞—Е –Є–Ј –љ–µ—А–ґ–∞–≤–µ—О—Й–µ–є —Б—В–∞–ї—М–љ–Њ–є –њ—А–Њ–≤–Њ–ї–Њ–Ї–Є –≤ –Ї–Њ–Љ–љ–∞—В–µ —Б –Ї–Њ–љ–і–Є—Ж–Є–Њ–љ–µ—А–Њ–Љ (22¬∞C) –Є 12-—З–∞—Б–Њ–≤—Л–Љ —Ж–Є–Ї–ї–Њ–Љ —Б–≤–µ—В–∞ –Є —В–µ–Љ–љ–Њ—В—Л. –Ю–љ–Є –Є–Љ–µ–ї–Є —Б–≤–Њ–±–Њ–і–љ—Л–є –і–Њ—Б—В—Г–њ –Ї –≤–Њ–і–µ –Є –њ–Є—В–∞–љ–Є—О —Б–Њ–≥–ї–∞—Б–љ–Њ –і–Є–µ—В–µ.
–Я–Њ–ї—Г—З–µ–љ–Є–µ —Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤
–Ъ—А—Л—Б –ї–Є—И–∞–ї–Є –њ–Є—Й–Є –љ–∞ 14 вАУ 16 —З–∞—Б–Њ–≤ –Є –њ–Њ—Б–ї–µ –≤–≤–µ–і–µ–љ–Є—П –≤ —Н—Д–Є—А–љ—Л–є –љ–∞—А–Ї–Њ–Ј –Њ—В–±–Є—А–∞–ї–Є –Ї—А–Њ–≤—М –≤ –њ—А–Њ–±–Є—А–Ї—Г, —Б–Њ–і–µ—А–ґ–∞—Й—Г—О –≠–Ф–Ґ–Ъ, –њ—Г—В–µ–Љ —Б–µ—А–і–µ—З–љ–Њ–є –њ—Г–љ–Ї—Ж–Є–Є. –Я–ї–∞–Ј–Љ—Г –Ї—А–Њ–≤–Є –Њ—В–і–µ–ї—П–ї–Є —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А–Њ–≤–∞–љ–Є–µ–Љ –њ—А–Є 1000 g –≤ —В–µ—З–µ–љ–Є–µ 5 –Љ–Є–љ. –≠—А–Є—В—А–Њ—Ж–Є—В—Л –њ–Њ–ї—Г—З–∞–ї–Є –њ–Њ—Б–ї–µ —Б–±–Њ—А–∞ –њ–ї–∞–Ј–Љ—Л.–Я–Њ—Б–ї–µ —Г–і–∞–ї–µ–љ–Є—П –ї–µ–є–Ї–Њ—Ж–Є—В–∞—А–љ–Њ–є –њ–ї–µ–љ–Ї–Є –Є –љ–µ–±–Њ–ї—М—И–Њ–є —З–∞—Б—В–Є –≤–µ—А—Е–љ–µ–≥–Њ —Б–ї–Њ—П —Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤, —Н—А–Є—В—А–Њ—Ж–Є—В—Л –њ—А–Њ–Љ—Л–≤–∞–ї–Є —В—А–Є —А–∞–Ј–∞ –≤ —Е–Њ–ї–Њ–і–љ–Њ–Љ –Є–Ј–Њ—В–Њ–љ–Є—З–µ—Б–Ї–Њ–Љ —Б–Њ–ї–µ–≤–Њ–Љ —А–∞—Б—В–≤–Њ—А–µ –њ—А–Є 1000 g –≤ —В–µ—З–µ–љ–Є–µ 5 –Љ–Є–љ –њ—А–Є 4ЋЪ–°. –Э–µ–±–Њ–ї—М—И—Г—О —З–∞—Б—В—М –≤–µ—А—Е–љ–µ–≥–Њ —Б–ї–Њ—П —Г–і–∞–ї—П–ї–Є –њ—А–Є –Ї–∞–ґ–і–Њ–є –њ—А–Њ–Љ—Л–≤–Ї–µ.
–С–Є–Њ—Е–Є–Љ–Є—З–µ—Б–Ї–Є–є –∞–љ–∞–ї–Є–Ј –њ–ї–∞–Ј–Љ—Л
–І–∞—Б—В—М –њ–ї–∞–Ј–Љ—Л –±—Л–ї–∞ –љ–∞–њ—А–∞–≤–ї–µ–љ–∞ –≤ –Ї–ї–Є–љ–Є—З–µ—Б–Ї—Г—О –ї–∞–±–Њ—А–∞—В–Њ—А–љ–Є—О (CRC Co, –Ю—Б–∞–Ї–∞, –ѓ–њ–Њ–љ–Є—П) –і–ї—П –њ—А–Њ–≤–µ–і–µ–љ–Є—П —Б—В–∞–љ–і–∞—А—В–љ—Л—Е –Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є—Е –ї–∞–±–Њ—А–∞—В–Њ—А–љ—Л—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є –њ–ї–∞–Ј–Љ—Л.–Т —З–∞—Б—В–љ–Њ—Б—В–Є, —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–љ—Л–є —Б–Њ—Б—В–∞–≤ –њ–ї–∞–Ј–Љ—Л –±—Л–ї —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ —Б –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–µ–Љ –∞–≤—В–Њ–Љ–∞—В–Є–Ј–Є—А–Њ–≤–∞–љ–љ–Њ–≥–Њ –∞–љ–∞–ї–Є–Ј–∞—В–Њ—А–∞ (Olympus AU-5200) –і–ї—П –Њ–њ—А–µ–і–µ–ї–µ–љ–Є—П –Њ–±—Й–µ–≥–Њ —Б–Њ–і–µ—А–ґ–∞–љ–Є—П —Е–Њ–ї–µ—Б—В–µ—А–Є–љ–∞, —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤ –Є —В—А–Є–∞—Ж–Є–ї–≥–ї–Є—Ж–µ—А–Њ–ї–∞ —Б–Њ–Њ—В–≤–µ—В—Б—В–≤—Г—О—Й–Є–Љ —Д–µ—А–Љ–µ–љ—В–∞—В–Є–≤–љ—Л–Љ —Б–њ–Њ—Б–Њ–±–Њ–Љ (—В–µ—Б—В—Л Cholsterol-C, Phospholipid-–Т –Є Triglyceride-G, Wako Pure Chemical Co, –Ю—Б–∞–Ї–∞, –ѓ–њ–Њ–љ–Є—П).
–Я–Њ–ї—Г—З–µ–љ–Є–µ –Њ–±—Й–Є—Е –ї–Є–њ–Є–і–Њ–≤
–≠–Ї—Б—В—А–∞–Ї—Ж–Є—П –Њ–±—Й–Є—Е –ї–Є–њ–Є–і–Њ–≤ –Є–Ј —Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤ –±—Л–ї–∞ –њ—А–Њ–Є–Ј–≤–µ–і–µ–љ–∞ —Б—А–∞–Ј—Г –ґ–µ –њ–Њ—Б–ї–µ –њ–Њ–ї—Г—З–µ–љ–Є—П –Њ—В–Љ—Л—В—Л—Е —Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤ [8,23,24]. 500 –Љ–Ї–ї —Г–њ–ї–Њ—В–љ–µ–љ–љ—Л—Е —Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤ –њ–Њ–і–≤–µ—А–≥–ї–Є –≥–µ–Љ–Њ–ї–Є–Ј—Г –≤ —А–∞–≤–љ–Њ–Љ –Њ–±—К–µ–Љ–µ –≤–Њ–і—Л. –Ъ –ї–Є–Ј–∞—В—Г –і–Њ–±–∞–≤–Є–ї–Є 4 –Љ–ї –Љ–µ—В–∞–љ–Њ–ї–∞, –∞ –Ј–∞—В–µ–Љ —З–µ—А–µ–Ј 40 –Љ–Є–љ—Г—В 4 –Љ–ї —Е–ї–Њ—А–Њ—Д–Њ—А–Љ–∞. –Х—Й–µ —З–µ—А–µ–Ј 30 –Љ–Є–љ —Н–Ї—Б—В—А–∞–Ї—В –±—Л–ї —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А–Њ–≤–∞–љ, –∞ –Њ—Б—В–∞—В–Њ–Ї –њ–Њ–≤—В–Њ—А–љ–Њ —Н–Ї—Б—В—А–∞–≥–Є—А–Њ–≤–∞–љ 4 –Љ–ї —Б–Љ–µ—Б–Є –Љ–µ—В–∞–љ–Њ–ї–∞ –Є —Е–ї–Њ—А–Њ—Д–Њ—А–Љ–∞ (1:1). –Ю–±—К–µ–і–Є–љ–µ–љ–љ—Л–µ —Н–Ї—Б—В—А–∞–Ї—В—Л –±—Л–ї–Є –њ—А–Њ–Љ—Л—В—Л 10 –Љ–ї KCl 0,88% –і–ї—П –њ–Њ–ї—Г—З–µ–љ–Є—П –і–≤—Г—Е—Д–∞–Ј–љ–Њ–є —Б–Љ–µ—Б–Є. 2 –Љ–ї –љ–Є–ґ–љ–µ–є —Д–∞–Ј—Л –≤—Л—Б—Г—И–Є–ї–Є –≤ –њ–Њ—В–Њ–Ї–µ –≥–∞–Ј–Њ–Њ–±—А–∞–Ј–љ–Њ–≥–Њ –∞–Ј–Њ—В–∞. –Ю–±—Й–Є–µ –ї–Є–њ–Є–і—Л –Є–Ј –њ–ї–∞–Ј–Љ—Л (500 –Љ–Ї–ї) –±—Л–ї–Є –њ–Њ–ї—Г—З–µ–љ—Л —В–µ–Љ –ґ–µ —Б–њ–Њ—Б–Њ–±–Њ–Љ, —З—В–Њ –Є –Є–Ј —Н—А–Є—В—А–Њ—Ж–Є—В–Њ–≤. –Ь–µ—В–∞–љ–Њ–ї –Є —Е–ї–Њ—А–Њ—Д–Њ—А–Љ, –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–љ—Л–µ –њ—А–Є —Н–Ї—Б—В—А–∞–Ї—Ж–Є–Є –ї–Є–њ–Є–і–Њ–≤, —Б–Њ–і–µ—А–ґ–∞–ї–Є –±—Г—В–Є–ї–≥–Є–і—А–Њ–Ї—Б–Є—В–Њ–ї—Г–Њ–ї (50 –Љ–≥/–ї).–†–∞–Ј–і–µ–ї–µ–љ–Є–µ —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–љ—Л—Е –Ї–ї–∞—Б—Б–Њ–≤
–†–∞–Ј–і–µ–ї–µ–љ–Є–µ —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–љ—Л—Е –Ї–ї–∞—Б—Б–Њ–≤, –≤–Ї–ї—О—З–∞—П –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л (Pls), –±—Л–ї–Њ –њ—А–Њ–≤–µ–і–µ–љ–Њ –њ–Њ –Њ–њ–Є—Б–∞–љ–љ–Њ–Љ—Г –љ–∞–Љ–Є –Љ–µ—В–Њ–і—Г [8]. –С—Л–ї–∞ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–∞ —Б–Є—Б—В–µ–Љ–∞ –Т–≠–Ц–• Agilent HPLC (HP-1100 Series, Agilent Technologies, –Ґ–Њ–Ї–Є–Њ, –ѓ–њ–Њ–љ–Є—П) —Б –Є—Б–њ–∞—А–Є—В–µ–ї—М–љ—Л–Љ –љ–µ—Д–µ–ї–Њ–Љ–µ—В—А–Є—З–µ—Б–Ї–Є–Љ –і–µ—В–µ–Ї—В–Њ—А–Њ–Љ, —Д–ї—Г–Њ—А–µ—Б—Ж–µ–љ—В–љ—Л–Љ –і–µ—В–µ–Ї—В–Њ—А–Њ–Љ –Є –£–§-–і–µ—В–µ–Ї—В–Њ—А–Њ–Љ. –°–Є—Б—В–µ–Љ–∞ –±—Л–ї–∞ –њ–Њ–і–Ї–ї—О—З–µ–љ–∞ –Ї —А–∞–±–Њ—З–µ–є —Б—В–∞–љ—Ж–Є–Є Chem Station (Agilent Technology), –Ї–Њ—В–Њ—А–∞—П –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–ї–∞—Б—М –і–ї—П –Ї–Њ–љ—В—А–Њ–ї—П –Є –∞–љ–∞–ї–Є–Ј–∞ —Е—А–Њ–Љ–∞—В–Њ–≥—А–∞–Љ–Љ. –Ю—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ–µ —Б–Њ–і–µ—А–ґ–∞–љ–Є–µ —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–љ—Л—Е –Ї–ї–∞—Б—Б–Њ–≤ –±—Л–ї–Њ –Є–Ј–Љ–µ—А–µ–љ–Њ –≤ –Ї–∞–ґ–і–Њ–є —Е—А–Њ–Љ–∞—В–Њ–≥—А–∞—Д–Є—З–µ—Б–Ї–Њ–є –Њ–±–ї–∞—Б—В–Є —Б –њ–Њ–Љ–Њ—Й—М—О –Є—Б–њ–∞—А–Є—В–µ–ї—М–љ–Њ–≥–Њ –љ–µ—Д–µ–ї–Њ–Љ–µ—В—А–Є—З–µ—Б–Ї–Њ–≥–Њ –і–µ—В–µ–Ї—В–Њ—А–∞.–Р–љ–∞–ї–Є–Ј –ґ–Є—А–љ–Њ–Ї–Є—Б–ї–Њ—В–љ–Њ–≥–Њ —Б–Њ—Б—В–∞–≤–∞ —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤
–Ф–∞–љ–љ—Л–µ –Њ –Ї–∞–ґ–і–Њ–Љ —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–µ –±—Л–ї–Є –њ–Њ–ї—Г—З–µ–љ—Л –Т–≠–Ц–• —Б –£–§-–і–µ—В–µ–Ї—В–Њ—А–Њ–Љ [23,24]. –†–µ–Ј—Г–ї—М—В–∞—В—Л –∞–љ–∞–ї–Є–Ј–∞ –ґ–Є—А–љ–Њ–Ї–Є—Б–ї–Њ—В–љ–Њ–≥–Њ —Б–Њ—Б—В–∞–≤–∞ —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤ —Б–Њ–Њ—В–≤–µ—В—Б—В–≤–Њ–≤–∞–ї–Є –љ–∞—И–Є—Е –њ—А–µ–і—Л–і—Г—Й–Є–Љ —А–∞–±–Њ—В–∞–Љ [23,24].–Р–љ–∞–ї–Є–Ј –і–∞–љ–љ—Л—Е
–Ь—Л –њ—А–Њ–∞–љ–∞–ї–Є–Ј–Є—А–Њ–≤–∞–ї–Є –і–∞–љ–љ—Л–µ —Б –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–µ–Љ –њ–∞—А–љ–Њ–≥–Њ (–і–≤—Г—Б—В–Њ—А–Њ–љ–љ–µ–≥–Њ) t-–Ї—А–Є—В–µ—А–Є—П —Б —Г—А–Њ–≤–љ–µ–Љ –Ј–љ–∞—З–Є–Љ–Њ—Б—В–Є —А <0,05.–Ъ–Њ–љ—Д–ї–Є–Ї—В –Є–љ—В–µ—А–µ—Б–Њ–≤
–Р–≤—В–Њ—А—Л –Ј–∞—П–≤–ї—П—О—В, —З—В–Њ —Г –љ–Є—Е –Њ—В—Б—Г—В—Б—В–≤—Г–µ—В –Ї–Њ–љ—Д–ї–Є–Ї—В –Є–љ—В–µ—А–µ—Б–Њ–≤.–Т–Ї–ї–∞–і –Ї–∞–ґ–і–Њ–≥–Њ –Є–Ј –∞–≤—В–Њ—А–Њ–≤
–°–Ь –њ—А–Њ–≤–µ–ї –≤—Б–µ —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В—Л –Є –њ–Њ–і–≥–Њ—В–Њ–≤–Є–ї —А—Г–Ї–Њ–њ–Є—Б—М —Б—В–∞—В—М–Є. –Ґ–Ъ –њ—А–Є–љ—П–ї —Г—З–∞—Б—В–Є–µ –≤ –Ї—А–Є—В–Є—З–µ—Б–Ї–Њ–Љ –Њ–±—Б—Г–ґ–і–µ–љ–Є–Є —А–µ–Ј—Г–ї—М—В–∞—В–Њ–≤. –Ъ–Ь –њ–Њ–ї—Г—З–Є–ї –Њ—З–Є—Й–µ–љ–љ—Л–µ –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ—Л. –Ґ–§ —Б–њ–ї–∞–љ–Є—А–Њ–≤–∞–ї –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–µ –Є —Г—З–∞—Б—В–≤–Њ–≤–∞–ї –≤ –Ї—А–Є—В–Є—З–µ—Б–Ї–Њ–Љ –Њ–±—Б—Г–ґ–і–µ–љ–Є–Є —А–µ–Ј—Г–ї—М—В–∞—В–Њ–≤. –Т—Б–µ –∞–≤—В–Њ—А—Л –њ—А–Њ—З–Є—В–∞–ї–Є –Є –Њ–і–Њ–±—А–Є–ї–Є –Њ–Ї–Њ–љ—З–∞—В–µ–ї—М–љ—Л–є –≤–∞—А–Є–∞–љ—В —А—Г–Ї–Њ–њ–Є—Б–Є.–С–ї–∞–≥–Њ–і–∞—А–љ–Њ—Б—В–Є
–≠—В–Њ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–µ –±—Л–ї–Њ –њ—А–Њ–≤–µ–і–µ–љ–Њ –±–ї–∞–≥–Њ–і–∞—А—П —З–∞—Б—В–Є—З–љ–Њ–Љ—Г —Д–Є–љ–∞–љ—Б–Є—А–Њ–≤–∞–љ–Є—О –Э–∞—Ж–Є–Њ–љ–∞–ї—М–љ–Њ–є –Є—Б—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М—Б–Ї–Њ–є –Њ—А–≥–∞–љ–Є–Ј–∞—Ж–Є–Є –ѓ–њ–Њ–љ–Є–Є –≤ –Њ–±–ї–∞—Б—В–Є —Б–µ–ї—М—Б–Ї–Њ–≥–Њ —Е–Њ–Ј—П–є—Б—В–≤–∞ –Є –њ–Є—В–∞–љ–Є—П.–°–≤–µ–і–µ–љ–Є—П –Њ–± –∞–≤—В–Њ—А–∞—Е
–Ш–љ—Б—В–Є—В—Г—В —А–µ–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–є —Д—Г–љ–Ї—Ж–Є–Є –њ–Є—В–∞–љ–Є—П, 2241 Kubara, Hisayama-chou, Kasuya-gun, Fukuoka 811-2501, Japan.–Ъ–∞—Д–µ–і—А–∞ –Є–љ—В–µ–≥—А–∞—В–Є–≤–љ–Њ–є —Д–Є–Ј–Є–Њ–ї–Њ–≥–Є–Є, –Т—Л—Б—И–∞—П —И–Ї–Њ–ї–∞ –Љ–µ–і–Є—Ж–Є–љ—Б–Ї–Є—Е –љ–∞—Г–Ї –£–љ–Є–≤–µ—А—Б–Є—В–µ—В–∞ –Ъ—О—Б—О, Fukuoka 812-8582, Japan.
–¶–µ–љ—В—А–∞–ї—М–љ—Л–є –Є—Б—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М—Б–Ї–Є–є –Є–љ—Б—В–Є—В—Г—В, Marudai Food Co. Ltd, Osaka, Japan.
–Ш–љ—Б—В–Є—В—Г—В —А–µ–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–є —Д—Г–љ–Ї—Ж–Є–Є –њ–Є—В–∞–љ–Є—П, 2241 Kubara, Hisayama-chou, Kasuya-gun, Fukuoka 811-2501, Japan.
–Я–Њ–ї—Г—З–µ–љ–Њ: 5 —Б–µ–љ—В—П–±—А—П 2012 –≥–Њ–і–∞
–Я—А–Є–љ—П—В–Њ: 18 –љ–Њ—П–±—А—П 2012 –≥–Њ–і–∞
–Ю–њ—Г–±–ї–Є–Ї–Њ–≤–∞–љ–Њ: 21 –љ–Њ—П–±—А—П 2012 –≥–Њ–і–∞
doi:10.1186/1476-511X-11-161
–§–Њ—А–Љ–∞ —Ж–Є—В–Є—А–Њ–≤–∞–љ–Є—П: Mawatari et al: Dietary plasmalogen increases erythrocyte membrane plasmalogen in rats. Lipids in Health and Disease 2012 11:161.
–Ф–∞–љ–љ—Л–є —В–µ–Ї—Б—В –Њ—Е—А–∞–љ—П–µ—В—Б—П –Ј–∞–Ї–Њ–љ–Њ–Љ –†–§ –Њ –Ј–∞—Й–Є—В–µ –∞–≤—В–Њ—А—Б–Ї–Є—Е –њ—А–∞–≤, –∞ —В–∞–Ї–ґ–µ –Љ–µ–ґ–і—Г–љ–∞—А–Њ–і–љ—Л–Љ –Ј–∞–Ї–Њ–љ–Њ–і–∞—В–µ–ї—М—Б—В–≤–Њ–Љ –≤ —Н—В–Њ–є –Њ–±–ї–∞—Б—В–Є.
–Ы—О–±–Њ–µ —З–∞—Б—В–Є—З–љ–Њ–µ –Є–ї–Є –њ–Њ–ї–љ–Њ–µ –Ї–Њ–њ–Є—А–Њ–≤–∞–љ–Є–µ –Є –≤–Њ—Б–њ—А–Њ–Є–Ј–≤–µ–і–µ–љ–Є–µ –±–µ–Ј —А–∞–Ј—А–µ—И–µ–љ–Є—П –≤–ї–∞–і–µ–ї—М—Ж–∞ –Ј–∞–њ—А–µ—Й–µ–љ–Њ. –Ы–Є—Ж–∞, –≤–Є–љ–Њ–≤–љ—Л–µ –≤ –љ–∞—А—Г—И–µ–љ–Є–Є –∞–≤—В–Њ—А—Б–Ї–Є—Е –њ—А–∞–≤ –Є –Є—Б–Ї–ї—О—З–Є—В–µ–ї—М–љ—Л—Е –њ—А–∞–≤ –љ–∞ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–µ —В–µ–Ї—Б—В–∞, –љ–µ—Б—Г—В –≥—А–∞–ґ–і–∞–љ—Б–Ї–Њ-–њ—А–∞–≤–Њ–≤—Г—О, –∞–і–Љ–Є–љ–Є—Б—В—А–∞—В–Є–≤–љ—Г—О –Є —Г–≥–Њ–ї–Њ–≤–љ—Г—О –Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ–Њ—Б—В—М"
–Э–∞—В—Г—А–∞–ї—М–љ—Л–є –Є—Б—В–Њ—З–љ–Є–Ї –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ –Є –Ї–Њ—Н–љ–Ј–Є–Љ–∞ Q10
Tamotsu
67 500 ₽
Tamotsu
67 500 ₽

Tamotsu
67 500 ₽
23
–Э–∞—В—Г—А–∞–ї—М–љ—Л–є –Є—Б—В–Њ—З–љ–Є–Ї –њ–ї–∞–Ј–Љ–∞–ї–Њ–≥–µ–љ–Њ–≤ –Є –Ї–Њ—Н–љ–Ј–Є–Љ–∞ Q10
